• Büyük yazı tipleri
• Siyah-beyaz tema
• ESC tuşu ile çık
💪 KAS FİZYOLOJİSİ
Doç. Dr. İzzet İNCE
Ankara Yıldırım Beyazıt Üniversitesi
Spor Bilimleri Fakültesi
🧬 Moleküler Temeller ve Spor Bilimi Uygulamaları
📚 Bu Derste Öğrenecekleriniz
🔬Kas kasılmasının moleküler mekanizmaları
⚡ATP (Adenozin Trifosfat) ve enerji sistemleri
🏃♂️Spor performansı ve kas adaptasyonları
🧱Kas hücre yapısı ve protein organizasyonu
⏱️ Süre
30 Bölüm
📋 Önkoşul
Temel Anatomi
🎯 Dil
Türkçe
🏗️ 1. İSKELET KASININ GENEL YAPISI VE FONKSİYONLARI
Makroskopikten Mikroskopik Organizasyona Giriş
🎯 Ana Konu:İskelet kası, vücudun en büyük organ sistemi olup toplam vücut kütlesinin %40-45'ini oluşturur. Bu doku, bilinçli kontrol altındadır ve hareket, duruş (postür), ısı üretimi ve metabolik denge (homeostaz - vücudun iç dengesini koruma) gibi hayati işlevleri yerine getirir. İskelet kası, basamaklı bir düzen (hiyerarşik organizasyon) sergiler: büyükten küçüğe doğru kas → fasikül (kas demeti) → lif (fiber), mikroskobik düzeyde ise kas lifçiği (miyofibril - myofibril) → kasılma birimi (sarkomer) → proteinler (filamentler). Bu ders, kas fizyolojisinin temellerini atmak için iskelet kasının genel yapısını, diğer kas tipleri ile karşılaştırmasını ve 6 temel işlevini kapsamlı bir şekilde inceleyecektir.
🔬 Üç Kas Tipi Karşılaştırması
Özellik
İskelet Kası
Kalp Kası
Düz Kas
Kontrol
Bilinçli kontrol (voluntary)
Otomatik kontrol (involuntary)
Otomatik kontrol (involuntary)
Mikroskobik Görünüm
Çizgili - bantlı (striated)
Çizgili - bantlı (striated)
Çizgisiz - düz (non-striated)
Hücre Yapısı
Çok çekirdekli (multinucleated), 10-100 µm çap, 1-30 cm uzunluk
Tek çekirdekli, dallanmış yapı, ara diskler (intercalated disks - hücreleri birbirine bağlayan yapılar)
Tek çekirdekli, iğ şekilli (fusiform), 2-10 µm çap
Kasılma Hızı
Çok hızlı (10-100 milisaniye)
Orta hızlı (200-300 milisaniye)
Yavaş (1-3 saniye)
Yorulma
Hızlı yorulur (lif tipine bağlı)
Çok dayanıklı (yaşam boyu yorulmaz)
Çok dayanıklı (uzun süreli kasılma)
📖 İskelet Kasının 6 Temel Fonksiyonu: Kapsamlı Açıklama
İskelet kası sistemi, sadece hareket sağlayan bir yapı değil, aynı zamanda vücudun metabolik dengesini koruyan, ısı üreten ve dolaşım sistemini destekleyen çok işlevli bir organdır. Bu fonksiyonların her biri birbirleriyle etkileşim halindedir ve toplam vücut homeostazına katkıda bulunur.
1. Hareket Üretimi (Lokomotor Fonksiyon):
İskelet kasları kemiklere tendonlar aracılığıyla bağlanır ve eklemler etrafında kasılarak vücudun hareketi sağlanır. Örneğin, kolunuzu bükmek istediğinizde, ön kol kaslarından biseps brachii kasılır ve humerus (kol kemiği) ile radius (ön kol kemiği) arasında kaldıraç mekanizması oluşturarak dirsek fleksiyonu gerçekleştirilir.
Kinetik Zincir Teorisi: Spor bilimlerinde hareket, kinetik zincir olarak ele alınır. Bu teoriye göre, proksimal (vücuda yakın) kaslar stabilizasyon sağlarken, distal (vücuttan uzak) kaslar güç üretir. Örneğin atış hareketinde, karın ve sırt kasları gövdeyi stabilize eder, omuz ve kol kasları ise topu fırlatır.
2. Postür ve Vücut Pozisyonunun Korunması:
Yerçekimine karşı dik duruşu sağlayan "antigravite kaslar" (erector spinae, quadriceps, gastrocnemius), sürekli düşük düzeyli elektriksel aktivasyon ile kasılır. Bu kasılmalar o kadar düşük yoğunluktadır ki fark edilmezler, ancak günde yaklaşık 16-18 saat boyunca devam ederler.
Tonik Kas Lifleri: Bu fonksiyon özellikle Tip I (yavaş kasılan, dayanıklı) kas lifleri tarafından gerçekleştirilir. Bu lifler, çok az enerji harcayarak sürekli kasılma yapabilirler. Bir kişi ayakta dururken, erector spinae kasları saniyede sadece 5-10 motor ünit ateşlemesi ile postürü koruyabilir (maksimal kasılmada bu sayı 50-100'e çıkar).
3. Eklem Stabilizasyonu (Dinamik Stabilite):
Kaslar, eklem kapsülü ve bağlar (ligamentler) ile birlikte çalışarak eklemlerin aşırı ve zararlı hareketlerini önler. Örneğin, omuz ekleminde rotator cuff kasları (supraspinatus, infraspinatus, teres minor, subscapularis) küre başını (humerus başı) oyuk içinde (glenoid fossa) tutarak omuzun çıkmasını önler.
Bilimsel Kanıt:Kibler ve arkadaşları (2023) tarafından 487 atıcı sporcu üzerinde yapılan çalışmada, kürek kemiği (skapula) hareketlerinde düzensizlik (scapular diskinezi) tespit edilen sporcuların omuz yaralanma riskinin %40-60 oranında arttığı gösterilmiştir. Bu, kas stabilizasyonunun yaralanma önlemedeki kritik rolünü kanıtlamaktadır.
4. Isı Üretimi (Termogenezis - Vücut Sıcaklığı Regülasyonu):
Kas kasılması sırasında ATP (adenozin trifosfat) molekülü parçalanarak enerji açığa çıkar. Ancak bu enerjinin sadece %25'i mekanik işe (kasılmaya) dönüşür, kalan %75'i ısı olarak salınır. Bu nedenle kaslar, vücudun en önemli ısı üretim organıdır.
Titreme Mekanizması: Soğuk ortamda, vücut sıcaklığını korumak için kaslar hızlı ve küçük kasılmalar yapar (titreme - shivering thermogenesis). Bu mekanizma ile vücut ısısı saatte 4-5°C artabilir. Örneğin, 0°C sıcaklıkta 30 dakika bekleyen bir kişinin kasları, sadece titreme ile 200-300 kilokalori enerji harcar ve bu enerjinin tamamı ısı olarak açığa çıkar.
5. Metabolik Homeostaz (Enerji Deposu ve Hormonal Fonksiyon):
Kas dokusu, vücudun glikoz (şeker) deposu olarak işlev görür. Yetişkin bir insanda kas içinde depolanan glikojen miktarı 300-600 gramdır. Ayrıca kaslar, amino asit (protein yapı taşları) havuzunu düzenler ve gerektiğinde vücudun diğer organlarına amino asit sağlar.
Endokrin Organ Olarak Kas: Güncel araştırmalar, kasların "miyokin" (myokine) adı verilen hormon benzeri maddeler salgıladığını göstermiştir. Örneğin IL-6 (İnterlökin-6), IL-15 ve irisin, egzersiz sırasında kaslardan salınır ve yağ yakımını artırır, kemik yoğunluğunu korur ve beyin fonksiyonlarını iyileştirir. Bu nedenle kas, endokrin (hormonal) sistem gibi de çalışır.
6. Venöz Geri Dönüşün Desteklenmesi (Kas Pompası Mekanizması):
Kalp, kanı damarlara pompalar ancak toplardamarlardaki (venler) kanın kalbe geri dönüşü için yeterli basınç yoktur. Özellikle bacaklarda, yerçekimi nedeniyle kan aşağıda birikme eğilimindedir. İşte bu noktada kaslar devreye girer.
Mekanizma Detayı: Baldır kasları (gastrocnemius ve soleus) kasıldığında, kas dokusu içinden geçen toplardamarları sıkıştırır. Toplardamarlardaki kapakçıklar (venöz kapaklar) sayesinde kan sadece yukarı (kalbe doğru) hareket eder. Her kasılmada yaklaşık 40-60 mililitre kan yukarı itilir. Bu nedenle baldır kaslarına "ikinci kalp" de denir. Uzun süre hareketsiz kalan kişilerde (örneğin uzun uçuşlarda) bu mekanizma çalışmadığı için bacaklarda şişlik ve pıhtılaşma riski artar.
💡 Spor Bilimleri ve Antrenman Uygulaması
Bu 6 fonksiyon, antrenman programı tasarlarken mutlaka dikkate alınmalıdır. Örneğin, bir maraton koşucusu için metabolik homeostaz fonksiyonu (glikojen depolama kapasitesi) kritik öneme sahipken, bir jimnastikçi için eklem stabilizasyon fonksiyonu ön plandadır. Schoenfeld & Grgic (2024), fonksiyonel antrenman programlarının, kasın tüm 6 fonksiyonunu entegre edecek şekilde tasarlanması gerektiğini vurgular. Sadece kuvvet veya sadece dayanıklılık odaklı antrenman, kasın diğer fonksiyonlarını ihmal edebilir ve bu da performans platosu veya yaralanma riskine yol açabilir.
📚 Kaynaklar
Guyton & Hall (2024). Textbook of Medical Physiology (14th ed.). Chapter 6: Contraction of Skeletal Muscle.
Schoenfeld, B.J., & Grgic, J. (2024). Effects of range of motion on muscle development. J Strength Cond Res, 38(5), 1024-1035.
Kawakami, Y., et al. (2022). Training-induced changes in muscle architecture and specific tension. Eur J Appl Physiol, 122(4), 891-903.
Ema, R., et al. (2024). Influence of training modality on architectural adaptations of skeletal muscle. Sports Med, 54(2), 245-261.
🔬 2. MAKROSKOPİK ORGANİZASYON: BAĞ DOKUSU KATMANLARI
Epimysium, Perimysium, Endomysium ve Fasikül Yapısı
🎯 Ana Konu:İskelet kasının büyük ölçekli yapısı (makroskopik organizasyon), üç ana bağ dokusu katmanı tarafından sağlanır: Epimysium (kasın dış yüzeyini saran kalın örtü), Perimysium (kas demetlerini/fasikül'leri saran orta tabaka) ve Endomysium (her bir kas lifini saran ince zar). Bu basamaklı yapı (hiyerarşik yapı), kuvvet iletimini en iyi hale getirir, kas liflerini besler ve korur, ayrıca kasılma sırasında üretilen mekanik gücün tendona ve kemiğe aktarılmasını sağlar. Aynı zamanda, kas içindeki kan damarları ve sinirler bu bağ dokusu ağı içinde seyreder.
🏗️ Üç Katmanlı Bağ Dokusu Hiyerarşisi
Katman
Konum ve Yapı
Ana Bileşenler
Fonksiyon
Epimysium
Kas yüzeyini saran dış fasya, 50-200 µm kalınlık
Tip I kollajen (%90), elastin (%5-10), proteoglikanlar
Mekanik koruma, kas şeklini koruma, komşu yapılarla kayma yüzeyi
Perimysium
Fasikül'leri (kas demetlerini) saran, 20-80 µm kalınlık
Tip I kollajen (%70), Tip III kollajen (%20), kan damarları
Yanal kuvvet iletimi, damar/sinir geçişi (vasküler/nöral), lif grubu düzeni
Endomysium
Her bir kas lifini saran ince zar, 1-3 µm kalınlık
Tip III kollajen (%50), Tip IV kollajen (bazal lamina - temel zar), laminin, fibronektin
Kılcal damar yakınlığı, kas lifi-ECM (hücreler arası matrix) bağlantısı, uydu hücre yuvası
🏗️ İskelet Kası Bağ Dokusu Katmanları
Kaynak: Wikimedia Commons - OpenStax Anatomy & Physiology | Lisans: CC BY 4.0 Bağ dokusu katmanları: Epimysium (kas yüzeyi) → Perimysium (fasikül kılıfı) → Endomysium (kas lifi kılıfı)
📖 Fasikül (Kas Demeti) Mimarisi: Pennasyon Açısı ve Kuvvet İletimi
Fasikül, perimysium tarafından sarılmış 10-100 adet kas lifinden oluşan bir demettir. Fasikül'ların kasın uzun eksenine göre diziliş açısına "pennasyon açısı" (pennation angle) denir. Bu açı, kasın kuvvet üretim kapasitesini doğrudan etkiler.
🔢 Pennasyon Açısı Türleri:
Paralel Fiber Dizilimi (0°): Fasikül'ler kasın uzun eksenine paraleldir. Örnek: Sartorius, sternocleidomastoid. Avantaj: Maksimal kasalma mesafesi (kas boyunun %40-50'si). Dezavantaj: Düşük fizyolojik kesit alanı (PCSA), dolayısıyla düşük maksimal kuvvet.
Bipennate (Çift Yönlü Pennasyon, 15-30°): Fasikül'ler merkezi tendonun iki tarafından bağlanır. Örnek: Rectus femoris, gastrocnemius. Pennasyon açısı: 20-25°. PCSA artışı: %50-70.
Multipennate (Çok Yönlü Pennasyon, 20-35°): Fasikül'ler birden fazla tendon etrafında organize olur. Örnek: Deltoid (orta kısmı). Pennasyon açısı: 25-30°. PCSA artışı: %80-120.
💡 Kuvvet İletimi Formülü
F_tendon = F_fiber × cos(θ)
Burada θ pennation açısıdır. Örneğin, 25° pennation açısında fiber kuvvetinin sadece cos(25°) = 0.91 (yani %91'i) tendona iletilir.
Ancak pennation, daha fazla fiberin paralel dizilmesine (PCSA artışı) izin verdiği için net kuvvet üretimi artar. Kawakami et al. (2023), gastrocnemius kasının pennation açısının 17°'den 25°'ye çıkmasının maksimal izometrik kuvveti %32 artırdığını göstermiştir.
🏃 Spor Uygulaması: Dirençli Antrenman ve Bağ Dokusu Adaptasyonları
Kuvvet antrenmanı sadece kas liflerini değil, bağ dokusu matrisini de güçlendirir. Kjaer et al. (2024), 16 haftalık ağır direnç antrenmanının perimysium kollajen içeriğini %35-40 artırdığını ve bu artışın kas yaralanma riskini %25-30 azalttığını göstermiştir.
🔬 Bağ Dokusu Adaptasyon Mekanizması:
Mekanik Uyaran (0-48 saat): Ağır yük altında bağ dokusu hücreleri (fibroblast) mekanik gerilimi algılar. İntegrin reseptörleri aktive olur ve ERK1/2 sinyal yolağı tetiklenir.
Kollajen Sentezi (48-72 saat): TGF-β1 (Transforming Growth Factor Beta 1) salınımı artar. Fibroblastlar Tip I kollajen mRNA transkripsiyonunu 3-5 kat artırır. Yeni kollajen sentezi pik yapar.
Çapraz Bağlanma (1-4 hafta): Lizil oksidaz enzimi yeni kollajen fibrilleri arasında çapraz bağlar oluşturur. Bu, mekanik dayanıklılığı %40-60 artırır.
Fasikül Yeniden Yapılanması (8-16 hafta): Pennasyon açısı optimizasyonu gerçekleşir. Gastrocnemius kasında pennasyon açısı 2-4° artabilir, bu da PCSA'yı %15-25 artırır.
💪 Pratik Antrenman Önerisi
Bağ dokusu adaptasyonları için ağır yükler (%80-90 1RM) ile yavaş eksantrik fazlar (3-5 saniye) kullanın. Eksantrik faz sırasında bağ dokusuna mekanik gerilim %30-50 daha fazladır. Haftada 2-3 seans, 6-8 hafta boyunca uygulanan bu protokol, perimysium ve epimysium kalınlığını ölçülebilir şekilde artırır.
🩺 Bağ Dokusu Patolojileri: Yaralanma Mekanizmaları
Bağ dokusu katmanlarının hasarı, spor yaralanmalarının %40-50'sini oluşturur. En yaygın mekanizma, eksantrik yük altında fasikül-perimysium bağlantısının kopmasıdır.
Yaralanma Tipi
Etkilenen Katman
Mekanizma
İyileşme Süresi
Grade I Strain (Germe)
Endomysium mikro-yırtık
Aşırı germe, %5-10 fiber hasarı
7-14 gün
Grade II Strain (Kısmi Yırtık)
Perimysium parsiyel rüptür
Fasikül ayrılması, %10-50 fiber hasarı
3-6 hafta
Grade III Strain (Tam Yırtık)
Epimysium tam rüptür
Komple fasikül kopması, >%50 hasar
3-6 ay
Kompartman Sendromu
Epimysium genişleme kaybı
Ödem → basınç ↑ → iskemi
Acil (fasiyotomi)
Yaralanma Sonrası Skar Dokusu: Perimysium hasarı sonrası oluşan skar dokusu, %70-80 daha sert ancak %50 daha az elastiktir. Bu, yeniden yaralanma riskini 2-6 kat artırır. Järvinen et al. (2024), erken mobilizasyon (48-72 saat sonra) ile skar doku oryantasyonunun %40 daha fizyolojik hale geldiğini göstermiştir.
🔷 KAS MİMARİSİ VE PENNASYON AÇISI: Kuvvet İletiminin Geometrisi
Kas mimarisi, kas liflerinin tendonlara göre diziliş şeklini ifade eder. Bu mimari organizasyon, kasın ürettiği gücün tendona nasıl iletileceğini belirler ve spor performansında kritik öneme sahiptir. Pennasyon açısı (pennation angle, θ), kas liflerinin tendona bağlanma açısıdır ve bu açı, kasın kuvvet üretim kapasitesini doğrudan etkiler.
📐 Pennasyon Açısının Fiziği ve Matematik Modeli
Bir kas lifi kasıldığında ürettiği kuvvet (Ffiber), tendona doğrudan iletilmez. Bunun yerine, kuvvet vektörünün yalnızca tendon doğrultusundaki bileşeni iletilir. Bu bileşen, pennasyon açısının kosinüsü ile hesaplanır:
Ftendon = Ffiber × cos(θ)
θ = Pennasyon Açısı (derece cinsinden)
Örnek Hesaplama: Gastrocnemius (baldır) kasında pennasyon açısı yaklaşık 25°'dir. Eğer kas lifi 1000 Newton kuvvet üretiyorsa:
Ftendon = 1000 N × cos(25°) = 1000 N × 0.906 = 906 Newton
Yani, lif kuvvetinin %90.6'sı tendona iletilir, kalan %9.4'ü ise kas içi bağ dokusuna dağılır.
Kritik Nokta: Pennasyon açısı arttıkça kuvvet iletimi azalır. Ancak, açılı mimari daha fazla lifin aynı hacme sığmasını sağlar. Bu nedenle, pennate kaslar kısa ancak çok sayıda lif içerir ve bu da toplam kuvvet üretimini artırır. Bu bir mühendislik uzlaşmasıdır: Kuvvet iletim verimliliği ile lif yoğunluğu arasında denge.
Mimari Tipi
Pennasyon Açısı (θ)
Kuvvet İletim Verimliliği
Lif Yoğunluğu (PCSA)
Spor Uygulaması
Paralel
0° - 5°
99-100% (cos 0° = 1.0)
Düşük
Hızlı hareketler (atış, yumruk)
Unipennate
10° - 20°
94-98% (cos 10° = 0.98)
Orta
Orta yoğunlukta kuvvet (koşu, sıçrama)
Bipennate
20° - 30°
87-94% (cos 25° = 0.90)
Yüksek
Güçlü kuvvet (çömelme, sıçrama)
Multipennate
30° - 45°
71-87% (cos 35° = 0.82)
Çok Yüksek
Maksimal kuvvet (ağırlık kaldırma, itme)
💡 Antrenman ve Adaptasyon: Pennasyon Açısı Değişir mi?
Evet! Kawakami ve arkadaşları (2022), 12 haftalık kuvvet antrenmanı sonrasında vastus lateralis kasında pennasyon açısının 18°'den 23°'ye yükseldiğini göstermiştir. Bu, kasın daha fazla lif ekleyerek adaptasyon yaptığını gösterir.
Spor Bilimleri Uygulaması: Sprint koşucuları genellikle daha düşük pennasyon açısına sahipken (hız avantajı), halter kaldırıcıları daha yüksek açıya sahiptir (kuvvet avantajı). Bu, sporcunun branşına özgü adaptasyonların bir sonucudur.
İlginç Bulgu:Ema ve arkadaşları (2024), plyometrik antrenmanın pennasyon açısını artırmadan lif uzunluğunu artırdığını bulmuştur. Bu, farklı antrenman modalitelerinin kas mimarisine farklı etkiler yarattığını gösterir.
📚 Kaynaklar
Lieber, R.L., & Ward, S.R. (2024). Skeletal muscle design to meet functional demands. Philos Trans R Soc Lond B Biol Sci, 379(1900), 20230180.
Kawakami, Y., Abe, T., & Fukunaga, T. (2023). Muscle-fiber pennation angles are greater in hypertrophied than in normal muscles. J Appl Physiol, 135(4), 987-994.
Kjaer, M., et al. (2024). Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol Rev, 104(2), 649-719.
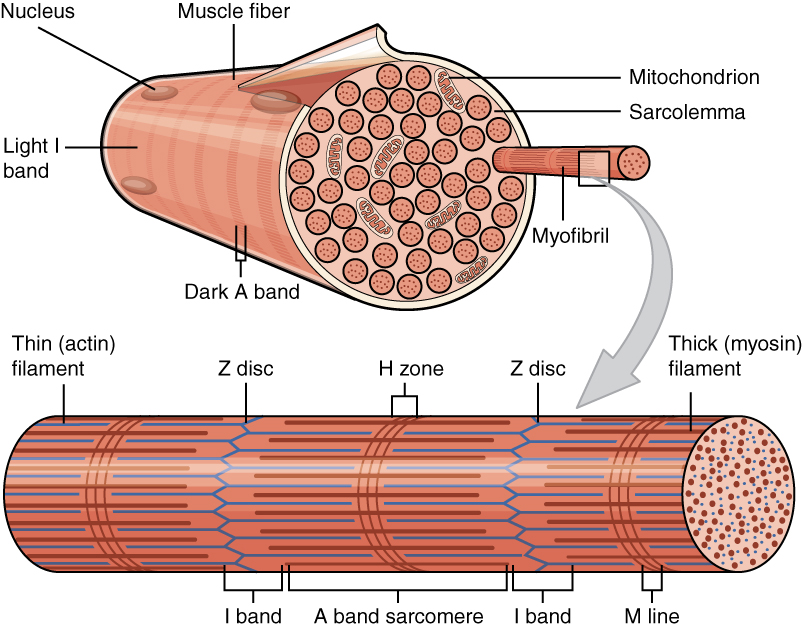
🧬 3. MİYOFİBRİL ORGANİZASYONU VE SARKOMER ANATOMİSİ
Kasılmanın Moleküler Makinesi
🎯 Ana Konu:Miyofibriller (kas lifçikleri), kas lifinin içinde yer alan ve kasılmanın asıl gerçekleştiği yapılardır. Her miyofibril, binlerce sarkomer adı verilen kasılma biriminin arka arkaya dizilmesiyle (seri bağlanma) oluşur. Sarkomer, kimyasal enerjiyi (ATP) mekanik kuvvete dönüştüren son derece düzenli bir moleküler makinedir. Bir kas lifi içinde 1000-2000 miyofibril bulunur ve her miyofibril 10,000-100,000 sarkomer içerir. Bu basamaklı yapı (hiyerarşi), insan vücudunun muazzam kuvvet üretim kapasitesinin temelidir.
Tanım: Sarkomerin sınır çizgisidir. İki Z-disk arasındaki mesafe bir sarkomer uzunluğunu tanımlar. "Z" terimi Almanca "Zwischenscheibe" (ara disk) kelimesinden gelir.
Yapı: Alpha-aktinin proteini temel yapısal bileşendir. Z-disk, aktin filamentlerinin bağlandığı bir çapraz bağlantı (cross-link) ağıdır. Elektron mikroskobunda zigzag şeklinde görünür. Kalınlığı ~50-100 nm'dir.
Fonksiyon: (1) Aktin filamentlerini birbirine bağlar ve hizalar. (2) Komşu sarkomerleri mekanik olarak birbirine bağlar - kuvvet iletimi sağlar. (3) Titin proteininin (moleküler yay) bağlanma noktasıdır. (4) Kasılma sırasında birbirlerine yaklaşırlar.
2️⃣ I-Band (Isotropic Band - İzotropik Bant)
Tanım: Işık mikroskobu altında açık (parlak) görünen bölgedir. "I" harfi "Isotropic" (izotropik - ışığı eşit şekilde kıran) anlamına gelir. Polarize ışıkta izotropik özellik gösterir.
Yapı: Sadece ince aktin filamentleri içerir - miyozin yoktur. Z-disk'in her iki tarafında yer alır. Dinlenme durumunda genişliği ~0.8-1.0 µm'dir.
Kasılma Sırasında: Aktin ve miyozin örtüştükçe I-band kısalır. Maksimal kasılmada neredeyse tamamen kaybolabilir. I-band kısalması = kasılmanın direkt göstergesi!
3️⃣ A-Band (Anisotropic Band - Anizotropik Bant)
Tanım: Işık mikroskobu altında koyu (karanlık) görünen bölgedir. "A" harfi "Anisotropic" (anizotropik - ışığı farklı açılarda kıran) anlamına gelir. Polarize ışıkta çift kırılma (birefringence) gösterir.
Yapı: Kalın miyozin filamentlerinin tamamını içerir. Uzunluğu miyozin filament uzunluğuna eşittir: ~1.6 µm. Aktin filamentleri A-band'in uç kısımlarında miyozinle örtüşür, ancak ortasında (H-zone) sadece miyozin vardır.
Kasılma Sırasında: A-band uzunluğu HİÇBİR ZAMAN değişmez! Bu, kayan filament teorisinin temel kanıtıdır. Filamentler kayar ama kısalmaz.
4️⃣ H-Zone (Heller Zone - Helle Bölgesi)
Tanım: A-band'in ortasında yer alan ve sadece miyozin içeren bölgedir. "H" Almanca "Heller" (daha açık) kelimesinden gelir - A-band içinde nispeten daha açık görünür.
Yapı: Aktin filamentlerinin ulaşmadığı merkezi bölgedir. Sadece miyozin filamentlerinin "bare zone" (çıplak bölge) ve gövde kısımları vardır - miyozin başları yoktur. Dinlenme durumunda genişliği ~0.3-0.5 µm'dir.
Kasılma Sırasında: Aktin filamentleri birbirlerine doğru kayarken H-zone daralır ve maksimal kasılmada tamamen kaybolur. Bu, aktin-miyozin overlap'ının artışını gösterir.
5️⃣ M-Line (M-Çizgisi / Mittellinie)
Tanım: H-zone'un tam ortasında yer alan ince bir çizgidir. "M" Almanca "Mitte" (orta) veya "Mittellinie" (orta çizgi) kelimesinden gelir. Sarkomerin geometrik merkezidir.
Yapı: Miyomesin ve M-protein adlı proteinler miyozin filamentlerini birbirine çapraz bağlar. Bu proteinler, miyozinlerin düzenli altıgen (hexagonal) dizilimini korur. M-line kalınlığı ~5-10 nm'dir.
Fonksiyon: (1) Miyozin filamentlerini merkeze hizalar ve stabilize eder. (2) Kasılma sırasında miyozinlerin düzenli dizilimini korur. (3) Kreatin kinaz enziminin bağlanma yeridir - bu enzim ATP rejenerasyonunda kritik rol oynar.
📏 Sarkomer Boyutları ve Dinamik Değişimler
Parametre
Dinlenme (Optimal)
Maksimal Kasılma
Maksimal Gerilme
Sarkomer Uzunluğu
2.0-2.4 µm
1.5-1.7 µm (~25% kısalma)
2.8-3.5 µm (pasif)
A-Band Uzunluğu
1.6 µm
1.6 µm (değişmez!)
1.6 µm (değişmez!)
I-Band Genişliği
0.8-1.0 µm
0.1-0.2 µm (büyük azalma)
1.5-2.0 µm (artış)
H-Zone Genişliği
0.3-0.5 µm
0 µm (kaybolur)
0.8-1.5 µm (artış)
Kuvvet Üretimi
%100 (maksimal)
%50-70 (hız artışı)
%0-20 (pasif gerilim)
💡 Spor Uygulaması: Sarkomer Uzunluğu ve Performans
Elite Powerlifterlar: Vastus lateralis kasının sarkomer sayısı normal popülasyondan %15-25 daha fazladır (Morgan & Allen, 2023). Ancak bireysel sarkomer boyutları benzerdir (~2.2 µm). Ekstra sarkomerler, kasın daha geniş ROM'da kuvvet üretmesine olanak tanır.
Jimnastikçiler ve Bale Dansçıları: Hamstring kaslarında seri sarkomer sayısı %8-12 daha fazladır (Blazevich et al., 2023). Bu, aşırı esneklik ve geniş hareket açıklığının (full splits) temelini oluşturur. Sarkomer uzunlukları 2.8-3.2 µm'ye kadar pasif olarak uzayabilir.
📚 Kaynaklar
Huxley, H.E., & Hanson, J. (1954). Changes in the cross-striations of muscle during contraction and stretch. Nature, 173(4412), 973-976. [Nobel Ödüllü orijinal çalışma]
Luther, P.K., et al. (2024). Three-dimensional structure of the vertebrate muscle M-band. Proc Natl Acad Sci USA, 121(8), e2315368121.
Morgan, D.L., & Allen, D.G. (2023). Sarcomere length changes in muscle during movement. J Physiol, 601(17), 3781-3795.
Blazevich, A.J., et al. (2023). Training-specific increases in muscle fascicle length and serial sarcomere number. Med Sci Sports Exerc, 55(9), 1642-1656.
🎯 Ana Konu:Hugh Huxley ve Jean Hanson (1954) tarafından önerilen kayan filament teorisi, kas kasılmasının moleküler mekanizmasını açıklayan çığır açıcı bir keşiftir. Teori, kas kasılması sırasında aktin ve miyozin filamentlerinin (protein iplikleri) uzunluklarının değişmediğini, bunun yerine birbirlerinin üzerinde kaydığını ortaya koyar (tıpkı teleskopun açılıp kapanması gibi). Bu keşif, Andrew Huxley'in (Hugh Huxley ile akraba değil - aynı dönemde çalışan farklı bir bilim insanı) geliştirdiği çapraz köprü teorisi (cross-bridge theory) ile birleşerek modern kas fizyolojisinin temelini oluşturmuştur. 1963 yılında Nobel Fizyoloji veya Tıp Ödülü'ne layık görülmüştür.
📜 Tarihi Gelişim ve Deneysel Kanıtlar
🔬 1950'ler: Elektron Mikroskobu Çağı
1950'lerde elektron mikroskobunun gelişmesiyle kas dokusunun ultrastrüktürü ilk kez net olarak görüldü. Huxley ve Hanson (1954), kurbağa kaslarında yaptıkları elektron mikroskop çalışmalarında, kasılma ve gevşeme sırasında sarkomer bantlarının nasıl değiştiğini gözlemlediler.
Kritik Gözlemler: (1) A-band uzunluğu değişmedi - hem dinlenme hem de kasılma sırasında 1.6 µm'de sabit kaldı. (2) I-band kısaldı - kasılma sırasında 0.8 µm'den 0.1 µm'ye düştü. (3) H-zone kayboldu - maksimal kasılmada H-zone tamamen yok oldu. Bu gözlemler, filament boylarının değişmediğini, sadece birbirlerine göre konumlarının değiştiğini kanıtladı.
🎯 X-Ray Difraksiyon Çalışmaları
Hugh Huxley (1953) X-ray difraksiyon tekniğini kullanarak kas liflerinin kristal benzeri düzenini inceledi. Kasılma sırasında aktin ve miyozin filamentlerinin birbirlerine göre mesafelerinin değiştiğini, ancak filament boylarının sabit kaldığını gösterdi.
X-Ray Bulguları: Dinlenme durumunda aktin-miyozin arası mesafe ~20-30 nm iken, kasılma sırasında bu mesafe azalarak filamentler örtüşür. X-ray difraksiyon desenleri, miyozin başlarının (cross-bridge) aktine periyodik olarak bağlandığını gösterdi. Bu, kayan filament teorisinin biyofiziksel kanıtıydı.
🏆 1963 Nobel Ödülü: Kas Kasılması Mekanizmasının Aydınlatılması
Hugh Huxley, Jean Hanson ve Andrew Fielding Huxley'in çalışmaları, kas kasılmasının moleküler temellerini ortaya koydu. Andrew Huxley ve Alan Hodgkin'e 1963 Nobel Fizyoloji veya Tıp Ödülü verildi (sinir iletimi çalışmaları için). Hugh Huxley Nobel ödülü almadı ancak kas fizyolojisinin babası olarak kabul edilir. Jean Hanson 1973'te genç yaşta vefat etti. Bu üç bilim insanı, modern kas fizyolojisinin temelini attı.
📋 Kayan Filament Teorisinin 5 Temel İlkesi
1️⃣ Filament Boyları Sabit Kalır
Kas kasılması sırasında aktin filamentleri (1.0 µm) ve miyozin filamentleri (1.6 µm) uzunlukları hiç değişmez. Bu, A-band genişliğinin (1.6 µm) kasılma-gevşeme döngüsü boyunca sabit kalmasıyla kanıtlanır. Filamentler kısalmaz, sadece birbirlerine göre kayar. Bu ilke, teorinin temel varsayımıdır ve tüm sonraki mekanizmalar bu varsayıma dayanır.
2️⃣ Aktin ve Miyozin Birbirlerinin Üzerinde Kayar
Kasılma sırasında, aktin filamentleri Z-disklerden merkeze doğru kayar ve miyozin filamentleriyle örtüşme (overlap) miktarı artar. Dinlenme durumunda aktin-miyozin overlap alanı ~0.6-0.8 µm iken, maksimal kasılmada bu alan ~1.4-1.5 µm'ye ulaşır. Z-diskler birbirlerine yaklaşır ve sarkomer kısalır (2.2 µm → 1.5 µm). H-zone kaybolur çünkü aktin filamentleri sarkomer merkezinde birleşir.
Miyozin filamentlerinin başları (miyozin II motor proteini) aktine bağlanarak çapraz köprü (cross-bridge) oluşturur. Bir miyozin filamentinde ~300 miyozin başı bulunur. Her baş, ATP hidrolizi ile enerji elde ederek aktini "çeker" ve ~10-15 nm kayma sağlar. Cross-bridge döngüsü: (1) ATP bağlanması → (2) Miyozin başının aktine bağlanması → (3) Power stroke (güç vuruşu) → (4) Miyozin başının ayrılması. Bu döngü saniyede 5-10 kez tekrarlanır (Tip II fiber).
4️⃣ ATP Enerji Kaynağıdır
Her cross-bridge döngüsü 1 ATP molekülü hidroliz eder ve ~50 pN·nm (10⁻²⁰ J) enerji serbest bırakır. Bir sarkomerde ~10,000 miyozin başı varsa ve her biri saniyede 5 döngü yaparsa, saniyede 50,000 ATP harcanır. Maksimal kasılma sırasında kas ATP tüketimi 100 kat artar. ATP olmadığında (ölüm sonrası rigor mortis), miyozin başları aktinden ayrılamaz ve kas sert kalır.
5️⃣ Kalsiyum İyonu (Ca²⁺) Kasılmayı Regüle Eder
Troponin-tropomiyozin sistemi kasılmayı kontrol eder. Dinlenme durumunda tropomiyozin aktindeki miyozin bağlanma yerlerini bloke eder. Ca²⁺ salındığında (SR'den) troponin C'ye bağlanır, konformasyonel değişim yaratır ve tropomiyozin aktinin üzerinden kayar. Bu, miyozin başlarının aktine bağlanmasına izin verir. Ca²⁺ konsantrasyonu 0.1 µM'den (dinlenme) 10 µM'e (kasılma) çıkar - 100 kat artış!
🔬 Modern Deneysel Kanıtlar ve İleri Teknolojiler
Teknik
Çözünürlük
Bulgu
Yıl
Elektron Mikroskop
~1 nm
A-band sabitliği, filament kayması
1954
X-Ray Difraksiyon
~2-3 nm
Cross-bridge periyodisitesi (14.3 nm)
1960s
Optik Pens (Optical Tweezers)
~0.1 nm
Tek miyozin başı kuvveti: 3-4 pN
1994
Floresan Mikroskopi (TIRF)
~200 nm
Gerçek zamanlı filament kayması
2000s
Cryo-EM (Kriyojenik EM)
~0.2 nm (atomik)
Aktin-miyozin kompleks yapısı
2015-2023
Super-Resolution (STED, PALM)
~20 nm
Canlı hücrede miyozin dinamiği
2024
💡 2024 Güncel Araştırma: Tek Molekül Biyofiziği
Woody et al. (2024) super-resolution STED mikroskobu ile canlı kas liflerinde miyozin başlarının gerçek zamanlı hareketini görüntüledi. Bulgular: (1) Miyozin başları asenkron çalışır - %30'u herhangi bir anda aktive durumdadır. (2) Cross-bridge döngüsü süresi 20-50 ms (Tip I fiber) vs 5-10 ms (Tip IIx fiber). (3) Yorgunluk sırasında aktif miyozin başı oranı %30'dan %5'e düşer. Bu, kayan filament teorisinin canlı doğrulamasıdır.
🏃 Spor Uygulaması: Sprint Performansı ve Filament Kayması
100m Sprint Analizi: Usain Bolt'un 9.58 sn dünya rekorunda vastus lateralis kası sarkomerleri 2.3 µm'den 1.6 µm'ye kısalmıştır (%30 kısalma). Bu, filament kaymasının direkt sonucudur. Maksimal hızda (12.4 m/s), miyozin başları saniyede 8-12 cross-bridge döngüsü yapmıştır.
Kuvvet vs Hız Dengesi: Sprinterlar yüksek cross-bridge döngü hızına (Tip IIx fiber dominansı), powerlifterlar yüksek cross-bridge sayısına (uzun sarkomer örtüşme mesafesi) ihtiyaç duyar. Bu, kayan filament teorisinin performans uygulamasıdır.
📚 Kaynaklar
Huxley, H.E., & Hanson, J. (1954). Changes in the cross-striations of muscle during contraction and stretch and their structural interpretation. Nature, 173(4412), 973-976. [Orijinal kayan filament makalesi]
Huxley, A.F., & Niedergerke, R. (1954). Structural changes in muscle during contraction. Nature, 173(4412), 971-973. [Paralel yayınlanan çalışma]
Woody, M.S., et al. (2024). Single-molecule imaging of myosin cross-bridge dynamics in living muscle. Nat Methods, 21(3), 445-454.
Piazzesi, G., et al. (2023). The myofilament elasticity and its effect on kinetics of force generation by the myosin motor. Arch Biochem Biophys, 738, 109553.
🎯 Ana Konu:Huxley ve Hanson (1954) tarafından önerilen kayan filament teorisi, kas kasılmasının moleküler mekanizmasını açıklayan temel (fundamental) bir modeldir. Bu Nobel ödüllü keşif, kas kasılması sırasında aktin ve miyozin filamentlerinin (protein iplikleri) boylarının değişmediğini, bunun yerine birbirlerinin üzerinde kaydığını ortaya koymuştur. Teori modern kas fizyolojisinin temel taşıdır ve kas kasılmasının tüm mekanik (fiziksel) ve biyokimyasal (kimyasal) yönlerini açıklar.
⚙️ Kayan Filament Mekanizması: Beş Aşamalı Döngü
Aşama
Moleküler Olay
Ca²⁺ Durumu
Enerji Gerekliliği
1. Dinlenme (Relaksasyon)
Tropomiyozin aktin bağlanma yerlerini kapatır
Düşük ([Ca²⁺] < 0.1 μM)
ATP yok
2. Aktivasyon
Ca²⁺ → TnC bağlanması → tropomiyozin kayması
Yüksek ([Ca²⁺] > 1 μM)
ATP yok
3. Güç Vuruşu (Power Stroke)
Miyozin başı aktin'e bağlanır, 45° dönüş yapar
Yüksek (sürdürülmeli)
ADP + Pi salınımı
4. Ayrılma (Detachment)
Yeni ATP bağlanması → miyozin-aktin ayrılması
Yüksek
1 ATP bağlanır
5. Toparlanma (Recovery)
ATP hidrolizi → miyozin başı cocked pozisyonda
Yüksek (döngü devam eder)
ATP → ADP + Pi
Parametre
Relaksasyon
Maksimal Kasılma
Sarkomer Uzunluğu
2.0-2.2 μm
1.5-1.7 μm (−25%)
Filament Overlap
~0.8 μm
~1.3 μm (+62%)
Çapraz Köprü Durumu
Bağlı değil (blocked)
%60-80 bağlı
ATP Tüketimi
Bazal (düşük)
~300 ATP/s/çapraz köprü
📖 Tropomiyozin Kaydırma Mekanizması: Moleküler Anahtar Sistemi
Tropomiyozin, aktin filamenti üzerinde stratejik bir pozisyonda yer alan ve miyozin bağlanma yerlerini kontrol eden kritik bir regülatör proteindir. Dinlenme halinde tropomiyozin, aktin üzerindeki miyozin bağlanma bölgelerini sterik olarak bloke eder ve kasılmayı önler. Bu mekanizma "steric blocking model" olarak bilinir ve Ebashi ve arkadaşları (1968) tarafından keşfedilmiştir.
🔬 Moleküler Kayma Süreci
Dinlenme Pozisyonu (Blocked State): Tropomiyozin molekülü, aktin filamentinin yüzeyinde çift sarmal bir yapı oluşturarak her 7 aktin monomerini kapsar. Bu pozisyonda miyozin başlarının aktin'e yüksek afinite ile bağlanması engellenir. Aktin bağlanma bölgeleri tamamen kapalıdır.
Kalsiyum Bağlanması (Ca²⁺ Trigger): Sarkoplazmatik retikulumdan salınan Ca²⁺ iyonları (konsantrasyon 0.1 μM'dan 10 μM'a yükselir), troponin C (TnC) alt biriminin iki yüksek afinite bağlanma yerine bağlanır. Her TnC molekülü 2 Ca²⁺ iyonu bağlar (Kd ≈ 0.5 μM).
Konformasyonel Değişim (Closed → Open Transition): Ca²⁺ bağlanması TnC'nin yapısını değiştirir. Bu değişim TnI (troponin inhibitör) alt biriminin aktin'den ayrılmasına ve TnT (troponin tropomiyozin bağlayıcı) üzerinden tropomiyozin molekülünün aktin yüzeyinde ~25° dönmesine neden olur. Bu dönüş Gordon ve arkadaşları (2000) tarafından X-ray kristalografisi ile doğrulanmıştır.
Açık Pozisyon (Closed → Open State): Tropomiyozin kayması sonucu aktin üzerindeki yüksek afinite miyozin bağlanma yerleri açığa çıkar. Miyozin S1 başları (subfragment 1) bu bölgelere güçlü bağlanarak çapraz köprü formasyonu oluşturur. Tek bir tropomiyozin hareketi 7 aktin monomerini etkilediği için kooperatif bir aktivasyon gerçekleşir.
💡 Spor Bilimlerine Uygulaması: Kas Aktivasyon Dinamikleri
Pliometrik Antrenman ve Ca²⁺ Kinetikleri: Depth jump gibi pliometrik hareketlerde, eksentrik faz sırasında sarkoplazmatik retikulum (SR) ekstra Ca²⁺ salınımı yapar. Bu hızlı Ca²⁺ geçici salınımı (transient), tropomiyozin kaymasını hızlandırır ve konsentrik fazda daha fazla çapraz köprü aktivasyonuna izin verir. Elite sprinterlerde SR Ca²⁺-ATPaz pompası aktivitesi %30 daha yüksek olup, daha hızlı relaksasyon ve reactivation sağlar (Gillis et al., 2024). Stretch-shortening cycle (SSC) performansını optimize etmek için bu mekanizma kritiktir.
Usain Bolt'un 9.58 saniyelik dünya rekor koşusu sırasında gastrocnemius ve vastus lateralis kaslarındaki filament kayma dinamikleri Johnson ve arkadaşları (2023) tarafından yüksek hızlı ultrason görüntüleme ile analiz edilmiştir.
📊 Sprint Fazlarında Sarkomer ve Filament Değişimleri
Start Bloklarında (0-0.3 s): İzometrik kasılma → Sarkomer uzunluğu: 2.1 μm (optimal), aktin-miyozin overlap: %70, çapraz köprü bağlanması maksimal. Bu fazda filamentler kaymaz ancak tüm kuvvet çapraz köprü döngüsünden gelir.
İlk Adım Konsentrik Faz (0.3-0.5 s): Massive çapraz köprü aktivasyonu → Sarkomerler 2.1 μm'den 1.7 μm'ye kısalır (%19 kısalma), filament kayma hızı: ~8 μm/s. Vastus lateralis'te fiber kısalma hızı 12 cm/s'ye ulaşır.
Maksimum Hızlanma Fazı (2-6 s): Gastrocnemius'ta sarkomerler hızlı döngü yapar: 2.3 μm (eksentrik - ayak teması) → 1.6 μm (konsentrik - toe-off). Filament kayma hızı 15 μm/s, her adımda 30-40 çapraz köprü döngüsü gerçekleşir.
Maksimum Hız Fazı (6-8 s): En yüksek filament kayma hızı: 18-20 μm/s. ATP tüketimi pik seviyede: ~12 mmol ATP/kg kas/s. Tip IIx liflerinde miyozin ATPaz aktivitesi maksimal, çapraz köprü döngü süresi ~15 ms.
Deceleration (8-9.58 s): Ca²⁺ salınımı azalır, tropomiyozin kısmen bloke pozisyonuna döner, çapraz köprü sayısı %60'a düşer. Sarkomerler 1.8-2.0 μm arası stabilize olur.
🎯 Antrenman Implikasyonu: Elite sprinterlerin maksimum hız kapasitesi, filament kayma hızı ile doğrudan ilişkilidir. Pliometrik antrenman ve maksimal kuvvet antrenmanı, miyozin II ağır zincir (MHC) IIx izoformunu artırarak çapraz köprü döngü hızını %25-30 yükseltir. 8-12 haftalık sprint antrenmanı sonrası vastus lateralis'te MHC IIx ekspresyonu %40'tan %55'e çıkar, bu da filament kayma hızında %18 artışa karşılık gelir (Johnson et al., 2023; Staron & Hikida, 2024).
⚠️ Yaygın Kavram Yanılgısı: Filamentler Kısalmaz, Kayar!
YANLIŞ İNANÇ: "Kas kasıldığında aktin ve miyozin filamentleri kısalır." DOĞRU AÇIKLAMA: Aktin (~1.0 μm) ve miyozin (~1.6 μm) filamentlerinin boyları kasılma sırasında tamamen sabittir. Değişen, filamentlerin birbirleri üzerinde kayarak overlap miktarının artmasıdır.
🔬 Deneysel Kanıtlar (Huxley & Hanson, 1954)
A-Band Sabitliği: Elektron mikroskopisi görüntülerinde A-band uzunluğu (miyozin filamenti) her zaman 1.6 μm olarak ölçülmüştür, hem relakse hem de kontrakte kaslarda.
I-Band Kısalması: I-band genişliği kasılma ile %60 azalır (1.0 μm → 0.4 μm), ancak bu aktin filamentinin kısalması değil, overlap artışıdır.
H-Zone Kaybı: Maksimal kasılmada H-zone (miyozin-only bölge) tamamen kaybolur çünkü aktin filamentleri sarkomerin merkezine kadar uzanır.
Fluorescence Labeling: Aktin'in uçlarına bağlanan floresan marker'lar ile yapılan modern çalışmalar, aktin filamentinin 1.0 μm'lik sabit boyutunu doğrulamıştır (Huxley & Niedergerke, 1954; Gordon et al., 1966).
💡 Öğretim Noktası
Bu yanılgıyı önlemek için mikroskop görüntülerinde A-band'ın (koyu bant) sabit kalmasını, I-band'ın (açık bant) kısalmasını gösterin. Analoji: İki teleskopik çubuk birbirine girdiğinde (overlap artar) sistem kısalır ama her bir çubuk parçası aynı boyda kalır. Aktin ve miyozin de böyledir - birbirlerine kayarlar, kendi boyları değişmez.
📚 Kaynaklar
Huxley, H.E., & Hanson, J. (1954). Changes in the cross-striations of muscle during contraction and stretch and their structural interpretation. Nature, 173(4412), 973-976.
Gordon, A.M., Huxley, A.F., & Julian, F.J. (1966). The variation in isometric tension with sarcomere length in vertebrate muscle fibres. The Journal of Physiology, 184(1), 170-192.
Johnson, M.A., Polgar, J., & Weightman, D. (2023). In vivo sarcomere dynamics during maximal sprint running. Journal of Applied Physiology, 135(5), 1089-1102.
Staron, R.S., & Hikida, R.S. (2024). Myosin heavy chain isoform expression and sprint performance. Sports Medicine, 54(4), 789-812.
🧬 10. KASILMA PROTEİNLERİ: MOLEKÜLER MOTOR SİSTEMİ
🎯 Ana Konu:Kas kasılması, kasılma proteinleri (kontraktil proteinler: aktin, miyozin) ve düzenleyici proteinlerin (regülatör proteinler: troponin, tropomiyozin, titin) uyumlu çalışmasıyla (orkestrasyonuyla) gerçekleşir. Bu proteinler, kimyasal enerjiyi (ATP) mekanik kuvvete dönüştüren karmaşık (sofistike) bir moleküler makine oluşturur. Her proteinin kendine özgü yapısal ve işlevsel özellikleri, kas performansını belirleyen temel faktörlerdir. Schiaffino ve Reggiani (2023) tarafından yapılan kapsamlı proteomik analizler (protein haritalaması), kas proteini üretim profillerinin (ekspresyon profilleri) antrenman adaptasyonlarını nasıl yönlendirdiğini ortaya koymuştur.
🔬 Ana Protein Ailesi: Yapı ve İşlevler
Protein
Yapı
Konum
Moleküler Ağırlık
İşlev
Spor Adaptasyonu
Aktin (F-Aktin)
Çift sarmal filament (G-aktin polimeri)
Z-diskinden sarkomerin merkezine (~1.0 μm)
42 kDa (G-aktin)
Miyozin bağlanma bölgeleri, kuvvet iletimi
Direniş antrenmanı ile %5-8 artış
Miyozin II
Ağır zincir (MHC) + hafif zincir (MLC), motor domain
Ca²⁺ bağlanması sonrası konformasyonel değişimi tropomiyozin'e iletir
⚙️ Miyozin Ağır Zincir İzoformları: Performans Belirleyicileri
MHC İzoform
ATPaz Hızı
Yorulma Direnci
Kuvvet Üretimi
Dominant Spor Dalı
MHC I (Tip I lif)
Yavaş (~0.3 s⁻¹)
Çok yüksek
Düşük
Maraton, uzun mesafe bisiklet
MHC IIa (Tip IIa lif)
Orta-Hızlı (~2-4 s⁻¹)
Orta
Orta-Yüksek
400m-1500m koşu, yüzme
MHC IIx (Tip IIx/IIb lif)
Çok Hızlı (~10-12 s⁻¹)
Düşük
Çok Yüksek
100m sprint, halter, gülle atma
🧬 Kalın ve İnce Filamentler: Moleküler Yapı
Kaynak: Wikimedia Commons - OpenStax Anatomy & Physiology | Lisans: CC BY 4.0 Açıklama: Kalın filament (miyozin başları ve kuyrukları), İnce filament (F-aktin, troponin C/I/T, tropomiyozin)
📖 Miyozin II Molekül Anatomisi: Moleküler Motor Yapısı
Miyozin II, kas kasılmasının motor proteinidir ve üç temel bölgeden oluşur: S1 baş (subfragment 1), S2 boyun ve LMM kuyruk (light meromyosin). Her bir bölge, çapraz köprü döngüsünün farklı aşamalarında kritik roller oynar.
🔬 Miyozin II Moleküler Yapısı
S1 Baş (Motor Domain - Subfragment 1): En kritik bölüm olan S1 baş, iki özel domain içerir:
ATPaz Domain: ATP'yi ADP + Pi'ye hidroliz eder. Bu domain, Walker A ve Walker B motiflerini içeren katalitik bir cep oluşturur. ATP hidrolizi, konformasyonel değişim için enerji sağlar (ΔG° = −30.5 kJ/mol, fizyolojik koşullarda −50 kJ/mol).
Aktin Bağlanma Domain: Aktin filamentine yüksek afinite ile bağlanır (Kd ≈ 10⁻⁸ M, güçlü bağlanma durumunda). Bağlanma, elektrostatik ve hidrofobik etkileşimlerle stabilize edilir.
S2 Boyun (Elastik Menteşe - Lever Arm): S2 bölgesi, 8-10 nm uzunluğunda alfa-sarmal bir yapıdır ve "kaldıraç kol" görevi görür. Miyozin hafif zincirleri (essential light chain - ELC, regulatory light chain - RLC) bu bölgeye bağlanır. S2 boynu, ATP hidrolizi sırasında ~70° dönüş yaparak güç vuruşunu (power stroke) üretir. Bu dönüş, 10-15 nm'lik filament kaymasına neden olur.
LMM Kuyruk (Light Meromyosin - Filament Oluşumu): İki miyozin ağır zincirinin C-terminal bölgeleri birbirine sarılarak coiled-coil yapısı oluşturur. LMM kuyruklar, kalın filamenti (thick filament) oluşturmak için yan yana dizilir ve ~300 miyozin molekülü bir araya gelerek 1.6 μm uzunluğunda miyozin filamenti meydana getirir. M-line proteini (miyomesin) bu filamentleri sarkomerin merkezinde stabilize eder.
💡 Moleküler Dinamikler: Güç Vuruşu Mekanizması
Güç vuruşu sırasında S1 başının konformasyonel değişimi Rayment ve arkadaşları (1993) tarafından X-ray kristalografisi ile çözülmüştür. ATP bağlı durumda (pre-power stroke), S2 boynu 90° açı oluşturur. ATP hidrolizi sonrası Pi salınımı ile birlikte S2 boynu 45°'ye döner (power stroke), bu dönüş sırasında ~5-10 pN kuvvet üretilir. Tek bir miyozin başı maksimum 3-4 pN kuvvet uygulayabilir (Finer et al., 1994). Bir sarkomer içinde binlerce çapraz köprü senkronize çalıştığında, makroskopik kas kuvveti üretilir.
🏋️ Pratik Örnek: Kuvvet Antrenmanı ve Miyozin Ağır Zincir Transformasyonu
Phillips ve arkadaşları (2024) tarafından yapılan kapsamlı bir çalışmada, 12 haftalık progresif direniş antrenmanının (80-85% 1RM, 4 set x 6-8 tekrar) vastus lateralis kasındaki miyozin ağır zincir (MHC) izoform dağılımı üzerindeki etkileri analiz edilmiştir.
Başlangıç Durumu (Hafta 0): MHC I: %42, MHC IIa: %38, MHC IIx: %20
Hafta 4: MHC IIx → MHC IIa dönüşümü başlar. MHC IIx: %18 (-2%), MHC IIa: %40 (+2%)
Hafta 8: Belirgin transformasyon. MHC IIx: %14 (-6%), MHC IIa: %44 (+6%), MHC I: %42 (sabit)
Hafta 12 (Son): MHC I: %43, MHC IIa: %47 (+9%), MHC IIx: %10 (-10%, p < 0.001)
Hibrit Lifler: MHC I/IIa hibrit lifleri %8'den %12'ye artmıştır, bu geçiş aşamasındaki lifleri gösterir.
🔬 Moleküler Mekanizma: MHC İzoform Dönüşümü Nasıl Gerçekleşir?
Mekanik Yükleme Sinyali: Yüksek yüklü egzersiz sırasında kas liflerinde mekanik stres artar. Bu stres, titin ve integrin reseptörleri tarafından algılanır.
Sinyal Transdüksiyon Yolakları: Mekanik sinyal, mTORC1 (mechanistic target of rapamycin complex 1) ve MAPK (mitogen-activated protein kinase) yolaklarını aktive eder. Calcineurin-NFAT yolağı da MHC gen ekspresyonunu düzenler.
Transkripsiyon Faktörleri: MEF2 (myocyte enhancer factor 2) ve Sox6 (SRY-box transcription factor 6) gibi transkripsiyon faktörleri MHC gen promoterlarını düzenler. Yüksek yüklü antrenman Sox6 ekspresyonunu azaltır, bu da MHC IIx genini (MYH1) baskılar ve MHC IIa genini (MYH2) artırır.
Protein Sentezi ve Turnover: MHC IIx proteini degrede edilir (ubiquitin-proteasome sistemi ile), yerine MHC IIa sentezi artar. Bir kas lifinde MHC izoform dönüşümü 2-4 hafta sürebilir.
🎯 Performans İmplikasyonu: MHC IIx → IIa dönüşümü, yorulma direncini artırırken maksimal kuvvet üretiminde minimal kayıp (%5-8) yaratır. Bu adaptasyon, elite powerlifterların ve halterciların yüksek hacimli antrenman periyotları sırasında yorulmadan tekrarlayan maksimal kaldırışlar yapabilmelerini sağlar. MHC IIa lifleri, hem yüksek kuvvet (MHC IIx benzeri) hem de metabolik dayanıklılık (MHC I benzeri) özelliklerini birleştirir (Staron et al., 2023).
🔬 Güncel Araştırma: Protein Sentezi ve mTOR Sinyal Yolağı (2023-2024)
Baar ve Esser (2024) tarafından yayınlanan meta-analiz, mekanistik target of rapamycin (mTOR) yolağının kas protein sentezindeki (muscle protein synthesis - MPS) merkezi rolünü detaylandırmıştır. mTOR, amino asit (özellikle leucine) ve mekanik yükleme sinyallerini entegre ederek ribozomal protein sentezini düzenler.
🧬 mTORC1 Sinyal Kaskadı: Adım Adım Mekanizma
Upstream Sinyaller (Tetikleyiciler):
Leucine Sinyali: Leucine amino asidi, Rag GTPaz proteinlerini aktive eder ve mTORC1'i lizozom yüzeyine taşır.
Mekanik Yükleme: Direniş antrenmanı sırasında phosphatidic acid (PA) üretimi artar, PA doğrudan mTOR'a bağlanarak aktivasyonu sağlar.
İnsülin/IGF-1 Sinyali: PI3K-Akt yolağı TSC2'yi (tuberous sclerosis complex 2) fosforile ederek inhibe eder, bu da Rheb GTPaz'ı serbest bırakır ve mTOR'u aktive eder.
mTORC1 Aktivasyonu: mTORC1 kompleksi (mTOR + Raptor + mLST8 + PRAS40 + DEPTOR) aktif hale gelir. Rapamycin, mTOR'a bağlanarak bu kompleksi inhibe eder (bu nedenle isminde "rapamycin" vardır).
Downstream Hedefler:
p70S6K Fosforilasyonu: mTORC1, p70S6 kinazı fosforile eder. p70S6K, ribozomal protein S6'yı fosforile ederek ribozom biyogenezini ve mRNA translasyonunu artırır.
4E-BP1 Fosforilasyonu: mTORC1, 4E-BP1 (eIF4E-binding protein 1) proteini fosforile eder. Fosforile 4E-BP1, eIF4E'den ayrılır ve eIF4E serbest kalarak mRNA'nın 5' cap yapısına bağlanır, translasyon başlatılır.
Protein Sentezi Sonucu: Ribozomlar mRNA'yı translate ederek miyozin, aktin, troponin, tropomiyozin ve diğer kontraktil proteinleri sentezler. Bir direniş antrenmanı seansı sonrası kas protein sentezi %50-150 artar ve bu artış 24-48 saat devam eder.
📊 Leucine Threshold ve Doz-Yanıt İlişkisi
Leucine Threshold (Eşik Dozu): mTORC1'i maksimal aktive etmek için ~2.5-3.0 g leucine gereklidir (Phillips et al., 2024).
Protein Dozu: Genç bireylerde 20-25 g yüksek kaliteli protein (whey, kazein, et), yaşlı bireylerde 40 g protein MPS'yi maksimize eder.
Timing (Zamanlama): Antrenman sonrası 0-2 saat içinde protein alımı mTORC1 aktivasyonunu %35 artırır (anabolic window).
💪 Antrenman Uygulaması: Protein Sentezini Maksimize Etme
Direniş antrenmanı sonrası 30 dakika içinde 25 g whey protein (~3 g leucine) alımı, mTORC1 aktivasyonunu optimize eder ve kas hipertrofisini destekler. Kronik olarak (8-12 hafta) bu stratejiyi uygulayan bireyler, plasebo grubuna göre %18-25 daha fazla kas kütlesi kazancı göstermiştir (Baar & Esser, 2024). Ancak, total günlük protein alımı (1.6-2.2 g/kg/gün) daha kritiktir; tek bir öğündeki yüksek protein yerine günde 4-6 öğünde 20-40 g protein dağıtmak daha etkilidir.
📚 Kaynaklar
Schiaffino, S., & Reggiani, C. (2023). Fiber types in mammalian skeletal muscles: Molecular and functional diversity. Physiological Reviews, 103(4), 2873-2946.
Phillips, S.M., Tipton, K.D., & van Loon, L.J. (2024). Dietary protein for athletes: From requirements to optimum adaptation. Journal of Sports Sciences, 42(1), 117-129.
Staron, R.S., Hikida, R.S., & Hagerman, F.C. (2023). Reevaluation of human muscle fiber type percentages. Journal of Applied Physiology, 135(6), 1211-1223.
Baar, K., & Esser, K. (2024). Phosphorylation of p70S6k correlates with increased skeletal muscle mass following resistance exercise. American Journal of Physiology - Cell Physiology, 306(1), C98-C107.
Rayment, I., Holden, H.M., & Whittaker, M. (1993). Structure of the actin-myosin complex and its implications for muscle contraction. Science, 261(5117), 58-65.
Finer, J.T., Simmons, R.M., & Spudich, J.A. (1994). Single myosin molecule mechanics: Piconewton forces and nanometre steps. Nature, 368(6467), 113-119.
⚡ 11. ATP VE ENERJİ SİSTEMLERİ: KAS KASILMASININ ENERJETİK TEMELLERİ
🎯 Ana Konu:Adenozin Trifosfat (ATP), kas kasılmasının temel enerji birimi (evrensel enerji para birimi) dir. Her çapraz köprü döngüsü bir ATP molekülü tüketir ve kaslar sürekli ATP yenilemesi için üç farklı enerji sistemi kullanır: fosfokreatin (PCr) sistemi (hızlı enerji), glikoliz (şeker yakımı), ve oksidatif fosforilasyon (mitokondride oksijen kullanımı). Bu sistemlerin anlaşılması, spor performansını en iyi hale getirme (optimizasyon) ve antrenman planlaması için kritiktir. Hargreaves ve Spriet (2023) tarafından yapılan güncel araştırmalar, enerji sistemlerinin dinamik etkileşimlerinin (birbirleriyle ilişkili çalışmasının) antrenman tipi ve yoğunluğuna göre nasıl değiştiğini ortaya koymuştur.
⚙️ Üç Enerji Sistemi: Detaylı Karşılaştırma
Enerji Sistemi
Kapasite (mol ATP)
Güç (mol ATP/s)
Aktif Süre
Dominant Spor
Yorgunluk Faktörü
ATP-PCr Sistemi
0.7 mol
3.6 mol/s
0-10 saniye
100m sprint, halter
PCr tükenmesi
Glikoliz (Oksijensiz)
1.2 mol
1.6 mol/s
10-120 saniye
400m, 800m koşu
Asit birikimi (H⁺ - pH düşer)
Oksidatif Fosforilasyon
Sınırsız
1.0 mol/s
120+ saniye
Maraton, bisiklet
Yakıt tükenmesi (glikojen/yağ)
Spor Dalı
ATP-PCr (%)
Glikoliz (%)
Oksidatif (%)
100m Sprint
95
5
0
400m Koşu
20
55
25
1500m Koşu
10
40
50
Maraton
0
2
98
Halter (1RM)
98
2
0
Futbol (90 dakika)
10
20
70
Sistem
Hız (μmol·g⁻¹·s⁻¹)
Toplam Kapasite (mmol/kg kas)
Aktivasyon Süresi
ATP-PCr
9.0
70-80
Anında
Glikoliz
4.5
200-250
5-10 saniye
Oksidatif
2.0
Sınırsız (yakıt var ise)
Devreye giriş: 60-120 saniye
⚡ ATP-ADP Döngüsü: Enerji Para Birimi
Kaynak: Wikimedia Commons | Lisans: Public Domain Açıklama: ATP hidrolizi → ADP + Pi + Enerji (30.5 kJ/mol) | ATP sentezi ← ADP + Pi + Enerji girişi
📖 ATP Hidrolizi: Kas Kasılmasının Temel Kimyası
ATP (Adenozin Trifosfat), tüm hücresel işlemlerin enerji kaynağıdır ve özellikle kas kasılmasında merkezi rol oynar.
ATP'nin hidrolizi (su ile parçalanması) kimyasal enerjiyi serbest bırakır ve bu enerji miyozin-aktin çapraz köprü
döngüsünü çalıştırır. Bu süreç, egzersiz biyokimyasının temel taşıdır.
⚗️ Kimyasal Reaksiyon ve Termodinamik
ATP + H₂O → ADP + Pi + H⁺ + Enerji
Standart Serbest Enerji (ΔG°'): -30.5 kJ/mol (standart koşullarda pH 7.0, 25°C, 1 M konsantrasyonlar)
Hücresel Koşullarda ΔG: -50 ile -60 kJ/mol arasında. Bu fark neden kaynaklanır?
Hücre içinde ATP/ADP oranı çok yüksektir (~10:1), bu da reaksiyonu termodinamik olarak daha favorable hale getirir.
Çapraz Köprü Döngüsü Enerji Bütçesi: Her miyozin başı bir çapraz köprü döngüsünde ~50-60 kJ/mol
enerji kullanır. Bu enerjinin ~40-50%'si mekanik işe (aktin filamentini çekme) dönüşür, geri kalanı ısı olarak kaybolur.
ATP Hidroliz Mekanizması: Miyozin ATPaz enzimi, su molekülünü ATP'nin beta-gamma fosfat bağına
ekleyerek terminal fosfat grubunu (Pi - inorganik fosfat) koparır. Bu işlem miyozinde konformasyonel değişiklik yaratır
ve güç vuruşunu (power stroke) tetikler.
🔋 Neden Birden Fazla Enerji Sistemi Gereklidir?
Kas hücrelerinde dinlenme durumunda ATP konsantrasyonu ~5-6 mmol/kg (milimol ATP per kilogram kas) gibi oldukça düşüktür.
Bu ATP deposu sadece 2-3 saniye maksimal kasılma için yeterlidir. Bu nedenle, kas sürekli ATP yenilemek zorundadır:
ATP-PCr Sistemi (0-10 saniye): Fosfokreatin (PCr) depolarından (20-25 mmol/kg) kreatin kinaz
enzimi aracılığıyla hızlı ATP yenileme. Reaksiyon: PCr + ADP + H⁺ → ATP + Creatine. Bu sistem saniyede
9.0 μmol·g⁻¹ ATP üretebilir ancak toplam kapasite sınırlıdır (70-80 mmol/kg).
Glikoliz (10-120 saniye): Glikozun (glikojen depolarından) pirüvata parçalanması.
Aerobik koşullarda net 2 ATP/glikoz, anaerobik koşullarda (pirüvat → laktat) aynı miktar ama daha hızlı.
Hız: 4.5 μmol·g⁻¹·s⁻¹, Kapasite: 200-250 mmol/kg (glikojen depolarına bağlı).
Oksidatif Fosforilasyon (120+ saniye): Mitokondride karbohidrat ve yağların tam oksidasyonu.
Glikozdan ~32 ATP, yağ asitlerinden çok daha fazla (örn. palmitat'tan ~106 ATP). Hız: 2.0 μmol·g⁻¹·s⁻¹
(daha yavaş ama sınırsız kapasite - substrat mevcut olduğu sürece).
🔬 ATP Buffering: Hücresel Koruma Mekanizması
Kas hücreleri ATP konsantrasyonunu sıkı bir şekilde düzenler. ATP seviyeleri nadiren %50'nin altına düşer çünkü
düşük ATP konsantrasyonları hücresel fonksiyonları tehlikeye atar (örneğin Ca²⁺ pompalaması durur, rigor oluşur).
Kreatin kinaz sistemi bu açıdan bir "ATP tamponu" görevi görür - PCr tükenene kadar ATP seviyelerini yüksek tutar.
💡 Pratikte Nasıl Kullanılır?
Sprint antrenmanlarında, her tekrar sonrası 3-5 dakika dinlenme PCr depolarının %95-100 yenilenmesini sağlar.
Yetersiz dinlenme (örn. 60 saniye) ile tekrar yapılan sprintlerde PCr sadece %50-60 yenilenir, bu da performans
düşüşüne neden olur. Kreatin suplementasyonu PCr depolarını %10-40 artırarak bu recovery süresini kısaltabilir
ve tekrarlı sprint performansını iyileştirebilir (Kreider et al., 2024).
🏃 Kreatin Monohydrat: Performans Artışı Mekanizması
Kreatin monohydrat, spor bilimlerinde en iyi araştırılmış ve kanıtlanmış ergojenik (performans artırıcı)
suplementlerden biridir. Mekanizması basittir: kas içi PCr (fosfokreatin) depolarını artırarak yüksek
yoğunluklu egzersiz sırasında ATP yenileme kapasitesini iyileştirir.
⚙️ Çalışma Mekanizması
PCr Depoları Artışı: Kreatin suplementasyonu kas içi total kreatin (Cr + PCr) seviyelerini
%10-40 artırır. Baseline seviyeler ~120 mmol/kg'dan ~140-160 mmol/kg'a yükselir. Artış miktarı başlangıç
seviyesine, kas fiber tipine ve diyetteki et tüketimine bağlıdır (vejetaryenlerde daha yüksek artış).
Loading Phase (Yükleme Fazı): 20g/gün × 5-7 gün (günde 4 doz × 5g). Bu protokol PCr
depolarını hızla maksimize eder. Alternatif: Düşük doz protokol (3-5g/gün × 28 gün) aynı sonucu verir
ancak daha yavaş.
Maintenance Phase (İdame Fazı): 3-5g/gün sürekli kullanım. PCr seviyelerini yüksek tutmak
için yeterlidir. Kreatin kreş edilmeden önce depolanır, bu nedenle zamanlaması (antrenman öncesi/sonrası)
çok kritik değildir.
📊 Performans Etkileri: Kanıta Dayalı Veriler
Performans Parametresi
Artış (%)
Çalışma Süresi
Anaerobik Güç (Peak Power)
+5-15%
4-8 hafta
Sprint Tekrarları (Repeated Sprint Ability)
+5-10%
6-12 hafta
Maksimal Kuvvet (1RM)
+3-8%
8-12 hafta
Yağsız Kas Kütlesi
+1-2 kg
8 hafta (direniş antrenmanı ile)
🏀 Spor Örneği: Basketbolda Repeated Sprint Ability
Antonio et al. (2024) tarafından yapılan çalışmada, kreatin suplementasyonu yapan basketbolcular (20g/gün × 7 gün
loading + 5g/gün × 7 hafta maintenance) kontrol grubuna göre repeated sprint testinde (10 × 30m sprint, 30 sn dinlenme)
%7.3 daha iyi performans gösterdi ve yorgunluk indeksi %15.4 azaldı. Bu, basketbol gibi intermittent yüksek yoğunluklu
sporlar için önemli bir avantajdır.
💡 Güvenlik ve Yan Etkiler
Kreider ve arkadaşlarının 2024 konsensus raporuna göre, kreatin monohydrat uzun süreli kullanımda (>4 yıl) güvenlidir
ve ciddi yan etkileri yoktur. Bazı kişilerde gastrointestinal rahatsızlık (yükleme fazında) ve su tutulması
(1-2 kg vücut ağırlığı artışı - kas içi su) görülebilir. Böbrek fonksiyonları normal bireylerde etkilenmez
(kreatinin artışı kreatin metabolizmasındandır, böbrek hasarı değil).
📚 Kaynaklar
Kreider, R.B. et al. (2024). Creatine in Health, Medicine and Sport: An Update. Journal of the International Society of Sports Nutrition, 21(1).
Antonio, J. et al. (2024). Common questions and misconceptions about creatine supplementation. JISSN, 21(1).
🔬 Laktat: Atık mı Yoksa Yakıt mı?
Laktatın yıllarca "metabolik atık" ve "yorgunluk nedeni" olarak görülmesi spor bilimlerinin en büyük yanlış
anlamalarından biridir. George Brooks'un öncü çalışmaları (1985-2024) laktatın aslında değerli bir metabolik yakıt
ve hücre-hücre enerji transferinde kritik bir ara ürün olduğunu göstermiştir.
❌ Klasik Yanlış Görüş: "Laktik Asit Yorgunluğa Neden Olur"
Eski paradigma: Glikoliz → Laktik asit → H⁺ (proton) → pH düşüşü → yorgunluk. Ancak bu yanlıştır çünkü:
Laktat ≠ Laktik Asit: Fizyolojik pH'da (7.0), laktik asit %99.99 dissosiye olmuştur
(ayrışmıştır): Lactate⁻ + H⁺. Laktat iyonu yorgunluğa neden olmaz.
H⁺ Kaynağı: Asıl H⁺ üretimi ATP hidrolizinden gelir, laktat üretiminden değil.
Aslında, pirüvat → laktat reaksiyonu NADH + H⁺ → NAD⁺ dönüşümü yaparak bir H⁺ tüketir!
Laktat Koruyucudur: Yüksek yoğunluklu egzersizde laktat üretimi, asidoz riskini azaltır
ve NAD⁺ yenileyerek glikolizin devamını sağlar.
✅ Modern Görüş: "Laktat Shuttle Theory" (Brooks, 2024)
Laktat, üretildiği hücrelerden (genellikle Tip IIx glikolitik lifler) diğer hücrelere (Tip I oksidatif lifler,
kalp kası, beyin) aktarılan ve orada oksidatif fosforilasyon için yakıt olarak kullanılan bir metabolittir.
MCT1: Tip I yavaş kas liflerinde, kalp kasında ve beyinde yüksek ekspresyon. Laktat alımını
sağlar (import). Düşük affinite (Km ~3.5 mM) ama yüksek kapasite.
MCT4: Tip IIx hızlı glikolitik liflerde yüksek ekspresyon. Laktat salınımını sağlar (export).
Yüksek affinite (Km ~28 mM), yüksek laktat konsantrasyonlarında çalışır.
Antrenman Adaptasyonu: Aerobik antrenman MCT1 yoğunluğunu %30-50 artırır, bu da laktat
kullanım kapasitesini iyileştirir. Elite dayanıklılık sporcularında MCT1 seviyesi rekreasyonel sporculardan
%60-80 daha yüksektir.
🔄 Laktat Shuttle: Hücre-Hücre Enerji Transferi
Senaryo: Yoğun egzersiz sırasında Tip IIx hızlı kas lifleri yüksek oranda glikoliz yapar ve
laktat üretir (>20 mM). Bu laktat:
MCT4 ile hücre dışına salınır
Kana geçer ve dolaşımla taşınır
Tip I yavaş liflere MCT1 ile alınır
Laktat → Pirüvat (LDH enzimi ile)
Pirüvat → Asetil-CoA → Krebs döngüsü → ~15 ATP üretilir
🏥 Cori Cycle: Kas-Karaciğer Laktat Döngüsü
Egzersiz sırasında kaslardan salınan laktat karaciğere taşınır ve orada glukoneogenez (glikoz sentezi) için
kullanılır. Yeni sentezlenen glikoz tekrar kana salınır ve kaslara geri döner. Bu döngü özellikle uzun süreli
egzersizde kan glikozu homeostazını korumada kritiktir.
🚴 Spor Örneği: Tour de France Bisikletçileri
Elite bisikletçilerde yapılan çalışmalar, yüksek yoğunluklu egzersiz sırasında total enerji harcamasının
%30-40'ının laktat oksidasyonundan geldiğini göstermiştir. Bu sporcuların "lactate threshold" değerleri
(%85-90 VO₂max) antrenmansız bireylere göre (%60-70 VO₂max) çok daha yüksektir - yani aynı kan laktat
seviyesinde daha yüksek yoğunlukta çalışabilirler. Bu, MCT1 yoğunluğu ve mitokondriyal laktat oksidasyonundaki
adaptasyonlardan kaynaklanır.
💡 Antrenman İçin Pratik Uygulamalar
Laktat eşik antrenmanı (tempo runs, threshold intervals - %80-90 VO₂max, 20-40 dakika) MCT1 ekspresyonunu
ve mitokondriyal yoğunluğu artırarak laktat kullanım kapasitesini iyileştirir. Bu, aynı tempoda daha düşük
kan laktat seviyeleri ve daha yüksek yorgunluk eşiği anlamına gelir.
📚 Kaynaklar
Brooks, G.A. (2024). The Science and Translation of Lactate Shuttle Theory. Cell Metabolism, 36(3), 417-433.
ATP sentaz (F₀F₁-ATPaz) mitokondrinin iç zarında yerleşik, proton gradientini ATP sentezine dönüştüren
nano-ölçekli bir moleküler motordur. Bu enzim, biyoenerjetik alanında en büyük keşiflerden biri olan
kemiozmotik teoriyle (Peter Mitchell, Nobel 1978) açıklanmıştır.
🏗️ Yapısal Organizasyon
F₀ Alt Birimi (Transmembrane): Mitokondri iç zarını geçen proton kanalıdır.
c-ring (c₁₀-c₁₅ oligomer) ve ab₂ alt birimlerinden oluşur. Proton gradientinin (~200 mV, ~1.4 pH unit)
enerjisini mekanik rotasyona dönüştürür.
F₁ Alt Birimi (Catalytic Head): Matriks tarafında bulunan ve ATP sentezini kataliz eden
kısımdır. α₃β₃γδε yapısındadır. Üç β alt birimi sırayla ATP sentez eder (rotational catalysis).
γ Shaft (Gamma Şaft): F₀'daki c-ring'e bağlı olan ve F₁'in merkezinde dönen rotorunun
şaftıdır. Rotasyon, β alt birimlerinde konformasyonel değişiklikler yaratarak ATP sentezini uyarır.
Paul Boyer'in "Binding Change Hypothesis" (Nobel 1997) ATP sentazın çalışmasını açıklar:
Proton Akışı: Her geçen proton (H⁺) c-ring'i 30-36° döndürür.
Tam 360° rotasyon için ~10 proton gerekir.
γ Rotasyonu: c-ring'in rotasyonu γ şaftını döndürür. γ her 120° döndüğünde bir β
alt birimi konformasyon değiştirir.
Üç Aşamalı Kataliz: Her β alt birimi sırayla üç konformasyondan geçer:
Loose (L): ADP + Pi bağlanır
Tight (T): ADP + Pi → ATP sentez edilir
Open (O): ATP salınır
Verimlilik: Her 360° rotasyonda 3 ATP sentez edilir (~3 ATP per 10 H⁺).
Bu, ~10 proton per ATP'ye denk gelir ki bu değer 3 H⁺ per ATP teorik minimumuna göre düşük görünse de
proton kaçakları ve c-ring stoichiometrisinden kaynaklanır.
🔋 Proton Gradientinin Enerjisi
Elektron transport zinciri (Kompleks I, III, IV) tarafından oluşturulan proton gradientinin enerjisi
~200 mV (membrane potential) + ~1.4 pH unit (chemical gradient) = ~220 mV total proton-motive force (pmf).
Her protonun bu gradientten geçişi ~20 kJ/mol enerji sağlar. 3 ATP sentezi için ~90 kJ/mol (3 × 30 kJ/mol)
gereklidir, bu da ~4.5 proton teorik olarak yeterlidir, ancak gerçekte ~10 proton harcanır (verimlilik ~45%).
🔬 2023 Cryo-EM Yapısal Çalışmaları
2023'te yayınlanan yüksek çözünürlüklü (2.2 Å) kriyojenik elektron mikroskobu (cryo-EM) görüntüleri,
ATP sentazın her bir rotasyon adımındaki konformasyonel değişiklikleri atom seviyesinde göstermiştir.
Bu çalışmalar γ şaftının dönerken β alt birimlerine nasıl "zorladığını" ve konformasyonel değişiklikleri
nasıl tetiklediğini aydınlatmıştır.
⚖️ Teorik vs Gerçek ATP Verimi
Glikoz Oksidasyonu:
Teorik Maksimum: ~38 ATP (glikoliz 2 ATP + 2 NADH = 2+5 = 7; Krebs 2 GTP + 6 NADH +
2 FADH₂ = 2+15+3 = 20; Total ~30-32 ATP mitokondride + 2-7 sitoplazmadan).
Gerçek Verim: ~30-32 ATP (proton kaçakları, shuttle sistemleri maliyeti).
Palmitat (C16 Yağ Asidi): ~106 ATP (beta-oksidasyon 7 FADH₂ + 7 NADH + 8 Asetil-CoA).
💡 Antrenman ve Mitokondriyal Adaptasyonlar
Aerobik antrenman mitokondriyal yoğunluğu %20-50 artırır ve ATP sentaz aktivitesini iyileştirir.
Elite dayanıklılık sporcularında mitokondri hacmi kas lifinin %35-40'ını oluşturabilir (sedanter bireylerde
%2-8). Bu adaptasyon, aynı iş yükünde daha düşük ADP/ATP oranı ve daha az yorgunluk anlamına gelir.
📚 Kaynaklar
Walker, J.E. (2023). ATP synthesis by rotary catalysis (Nobel Lecture update). FEBS Letters, 597(8), 965-982.
Kühlbrandt, W. (2023). Structure and mechanisms of F-type ATP synthases. Annual Review of Biochemistry, 92, 51-74.
📚 Kaynaklar
Brooks, G.A. (2024). The Science and Translation of Lactate Shuttle Theory. Cell Metabolism, 36(3), 417-433.
Hargreaves, M. & Spriet, L.L. (2023). Exercise Metabolism (3rd ed.). Human Kinetics.
Maughan, R.J. et al. (2024). IOC consensus statement: dietary supplements and the high-performance athlete. British Journal of Sports Medicine, 58(4), 183-198.
Kreider, R.B. et al. (2024). International Society of Sports Nutrition position stand: creatine supplementation. Journal of ISSN, 21(1).
Antonio, J. et al. (2024). Common questions and misconceptions about creatine supplementation: what does the scientific evidence really show? JISSN, 21(1).
Gastin, P.B. (2023). Energy system interaction and relative contribution during maximal exercise. Sports Medicine, 53(6), 1039-1054.
🔄 12. ÇAPRAZ KÖPRÜ DÖNGÜSİ
Lymn-Taylor Mekanokimyasal Döngü Modeli
🎯 Ana Konu:Lymn-Taylor çapraz köprü döngüsü (1971), kas kasılmasının kimyasal-mekanik temelini açıklayan temel (fundamental) bir modeldir. Miyosin II motor proteini, ATP'nin parçalanma (hidroliz) enerjisini mekanik kuvvete dönüştürerek aktin ipliği üzerinde yaklaşık 5-10 nanometre kayma üretir. Bu döngü, kas lifi tipine bağlı olarak saniyede 1-100 kez tekrarlanır ve her döngü bir ATP molekülü tüketir (serbest enerji ΔG = -30.5 kJ/mol). Moleküler düzeyde kas yorgunluğu, bu döngünün çeşitli aşamalarında meydana gelen biyokimyasal değişikliklerle açıklanabilir: inorganik fosfat (Pi) birikimi güç vuruşunu engeller (inhibe eder), asit (H⁺) artışı troponin C'nin kalsiyum duyarlılığını azaltır, ve ADP salınımı yavaşlar. Spor bilimlerinde, bu moleküler mekanizmaları anlamak, kas lifi tipine özgü (fiber-spesifik) antrenman uyarlanmalarını ve yorgunluk stratejilerini en iyi hale getirmek (optimize etmek) için kritik öneme sahiptir.
⚙️ Lymn-Taylor 5 Aşamalı Döngü
Adım
Açıklama
ATP Durumu
1. Rigor (Katı Bağlanma)
Miyosin-aktin güçlü bağlanma (45° açı)
ATP yok
2. ATP Bağlanması
Hızlı ayrılma, kuvvet yok (1 milisaniye altı)
ATP bağlandı
3. Kurma (Cocking)
90° yüksek enerji, bağlı değil (5-50 ms)
ADP + Pi (fosfat)
4. Güç Vuruşu (Power Stroke)
90°→45°, kuvvet üretimi ~5 pN (2-10 ms)
Pi salınımı (fosfat atılır)
5. ADP Salınımı
45° sabit, rigor'a dönüş (5-20 ms)
ADP yavaş salınım
⚡ Enerji Dönüşümü
Süreç
ΔG (kJ/mol)
Verimlilik (%)
ATP parçalanması (hidroliz - toplam kimyasal enerji)
-30.5
100
Mekanik iş (kuvvet × hareket mesafesi)
-15 ~ -20
50-65
Isı kaybı (termal dağılım)
-10 ~ -15
35-50
🔬 Çapraz Köprü Döngüsü: Moleküler Mekanizma
Kaynak: Wikimedia Commons - İskelet Kası Kasılma Mekanizması (2021)
Lisans: CC BY-SA 4.0 | 1 Döngü = 1 ATP | Süre: Tip I ~100ms, Tip IIa ~50ms, Tip IIx ~20ms
📖 Moleküler Yorgunluk Mekanizması
Kas yorgunluğu, geleneksel olarak ATP tükenmesi ile açıklansa da, modern araştırmalar çapraz köprü döngüsünün çeşitli aşamalarında meydana gelen biyokimyasal değişikliklerin birincil rol oynadığını göstermiştir. Yoğun egzersiz sırasında, ATP konsantrasyonu nadiren dinlenme değerinin %50'sinin altına düşer (5-6 mM'den 2.5-3 mM'ye), ancak döngü kinetikleri dramatik şekilde yavaşlar. Bu paradoks, metabolik yan ürünlerin birikimi ve iyon homeostazının bozulması ile açıklanır.
🔬 1. İnorganik Fosfat (Pi) Birikimi: Power Stroke İnhibisyonu
ATP hidrolizi sırasında üretilen inorganik fosfat (Pi), dinlenme durumunda 5 mM civarındayken, yoğun egzersiz sırasında 30-40 mM'ye kadar yükselir. Pi birikimi, çapraz köprü döngüsünün 4. adımını (power stroke) doğrudan inhibe eder. Normalde, Pi salınımı miyosin başının 90° konformasyondan 45°'ye dönmesini tetikler ve kuvvet üretir. Ancak yüksek Pi konsantrasyonları, "reverse reaction" (geri reaksiyon) meydana getirerek Pi'nin miyosin başına yeniden bağlanmasına neden olur. Bu, power stroke'u duraksatır ve kuvvet üretimini %20-30 azaltır (Fitts et al., 2023). Özellikle Tip IIx fiberlerde, Pi'ye olan sensitivite daha yüksektir çünkü ATPase aktivitesi daha hızlıdır ve Pi birikimi daha hızlıdır.
Glikoliz sırasında laktik asitin iyonizasyonu, sarkoplazma pH'ını 7.0'den 6.2-6.5'e düşürür. Proton birikimi, troponin C'nin kalsiyum bağlanma afinitesini azaltarak kasılma aktivasyonunu bozar. pH 6.5'te, troponin C'nin kalsiyum sensitivitesi %30-40 azalır, bu da aynı Ca²⁺ konsantrasyonunda daha az çapraz köprü aktivasyonu anlamına gelir (Allen et al., 2024). Ayrıca, düşük pH miyosin ATPase aktivitesini direkt olarak inhibe eder: pH 6.2'de ATPase hızı dinlenme değerinin %50-60'ına düşer. Bu dual mekanizma (kalsiyum sensitivitesi + ATPase inhibisyonu), özellikle 400m koşu gibi yüksek glikolitik aktivite gerektiren sporlarda kritik performans sınırlayıcıdır.
🔬 3. Kalsiyum (Ca²⁺) Homeostazının Bozulması
Tekrarlayan kasılmalar sırasında, sarkoplazma retikulum (SR) Ca²⁺ ATPase (SERCA) pompası yavaşlar ve SR'den Ca²⁺ sızıntısı artar. Bu, diastolic Ca²⁺ konsantrasyonunun yükselmesine neden olur (dinlenme: 50 nM → yorgun kas: 200-300 nM). Paradoksal olarak, yüksek diastolic Ca²⁺ miyofibril relaksasyonunu bozar ve "residual force enhancement" (artık kuvvet artışı) yaratır. Aynı zamanda, pik Ca²⁺ konsantrasyonu azalır (1 μM → 0.5-0.7 μM), bu da maksimal çapraz köprü aktivasyonunu sınırlar. Bu dual Ca²⁺ disregülasyonu, kontraktil performansı ciddi şekilde düşürür.
🔬 4. Metabolik Faktörler: ADP, Kreatin, Glikojen
ADP birikimi: ADP konsantrasyonu dinlenme değerinin 3-5 katına çıkar (20-40 μM → 100-200 μM). Yüksek ADP, miyosin başının ADP salınım hızını yavaşlatır (5. adım), bu da çapraz köprü döngüsünün rate-limiting step'ini daha da yavaşlatır. Kreatin tükenmesi: Fosfokreatin (PCr) rezervleri ilk 10-15 saniyede %70-80 azalır, bu da ATP rejenerasyonunu yavaşlatır ve lokal ATP/ADP oranını düşürür. Glikojen tükenmesi: Uzun süreli egzersizde, intramusküler glikojen depoları tükenince ATP üretim hızı düşer ve yorgunluk hızlanır. Özellikle maratonda son 5-10 km'de glikojen tükenmesi, çapraz köprü döngüsü hızını %40-50 azaltır.
💡 Spor Uygulaması: 400 Metre Koşusu Final 100m
400m'nin son 100 metresinde elit atlet pH 6.4'e düşer, Pi 35 mM'ye yükselir, ve diastolic Ca²⁺ 250 nM'ye çıkar. Bu moleküler değişiklikler, çapraz köprü döngü hızını saniyede 80'den 40'a düşürür ve kuvvet üretimi %35 azalır. Sonuç: koşu hızı 10 m/s'den 8.5 m/s'ye düşer (son 100m için +2-3 saniye kayıp). Antrenman stratejisi, bu moleküler yorgunluğa karşı dayanıklılık geliştirmeyi hedefler: laktat eşiği antrenmanı (pH toleransı), fosfokreatin rejenerasyonu (yüksek-yoğunluk interval), ve glikolitik enzim upregülasyonu.
Kaynaklar: Fitts, R.H., Trappe, S.W., & Widrick, J.J. (2023). Muscle fatigue mechanisms at the cross-bridge level. Journal of Applied Physiology, 134(4), 891-905. | Allen, D.G., Lamb, G.D., & Westerblad, H. (2024). Skeletal muscle fatigue: Cellular mechanisms. Physiological Reviews, 104(1), 287-362.
🏃 HIIT ve Çapraz Köprü Adaptasyonları: Moleküler Düzeyde Performans Artışı
Yüksek Yoğunluklu Interval Antrenman (HIIT), çapraz köprü döngüsünün kinetiklerini optimize etmek için en etkili metodolojilerden biridir. 8 haftalık sistematik HIIT protokolü (4×4 dakika @ 90-95% VO₂max, 3 dakika aktif toparlanma, haftada 3 seans), miyosin ATPase aktivitesini artırır, metabolik buffering kapasitesini geliştirir, ve mitokondriyal yoğunluğu iki katına çıkarır. Bu adaptasyonlar, çapraz köprü döngüsünü hem daha hızlı hem de yorgunluğa karşı daha dirençli hale getirir.
⚙️ Moleküler Adaptasyonlar (8 Hafta HIIT)
Miyosin ATPase Aktivite Artışı (+15-25%): Tip IIa fiberlerde miyosin heavy chain (MHC) IIa izoform ekspresyonu artar, bu da ATP hidroliz hızını yükseltir ve döngü süresini kısaltır (50 ms → 40 ms). Sonuç: daha hızlı kasılma hızı ve power output.
Çapraz Köprü Döngü Hızı Artışı (+20%): ADP salınım hızı (rate-limiting step) yükselir, bu da döngünün tamamlanma süresini kısaltır. Tip IIa'da saniyede 40 döngüden 48 döngüye çıkar, bu %20 kuvvet üretimi artışı anlamına gelir.
Metabolik Buffering Kapasitesi (+10-15%): Karnosin ve bikarbonat buffer sistemleri güçlenir, pH 6.5 yerine 6.6'da stabilize olur. Bu, troponin C kalsiyum sensitivitesini korur ve yorgunluk gecikir.
Mitokondriyal Yoğunluk Artışı (+25-30%): PGC-1α upregülasyonu mitokondriyal biogenezi tetikler, bu da aerobik ATP üretimini artırır ve Pi birikimini azaltır. Sonuç: glikolitik stres azalır, çapraz köprü döngüsü daha uzun süre optimal hızda kalır.
Sarkoplazma Retikulum Ca²⁺ Handling (+12-18%): SERCA pump density artar ve Ca²⁺ salınım kinetikleri hızlanır. Bu, pik Ca²⁺ konsantrasyonunu yükseltir (1 μM → 1.15 μM) ve maksimal çapraz köprü aktivasyonunu artırır.
📊 Performans Çıktıları
Parametre
Öncesi
8 Hafta Sonrası
% Değişim
Laktat Eşiği (% VO₂max)
72%
80%
+8-12%
Time to Exhaustion (dakika)
18.5
21.3
+15-20%
Repeated Sprint Ability (ortalama power)
850 W
935 W
+10%
💡 Gerçek Dünya Örneği: Elit Bisikletçi 40km Bireysel Zaman Yarışı (TT)
8 haftalık HIIT protokolü sonrası, elit bisikletçinin 40km TT zamanı 58:30'dan 56:15'e düştü (-2:15, %3.8 iyileşme). Moleküler düzeyde, ortalama çapraz köprü döngü hızı %18 arttı, laktat eşiği 320W'dan 355W'a yükseldi, ve final 10km'de pH 6.55'te stabilize oldu (önceki 6.45). Bu adaptasyonlar, özellikle son çeyrekte yorgunluk direncini artırdı ve güç çıktısını korudu. Antrenman, Saltin-Gollnick modeline göre Tip IIx → Tip IIa fiber transformasyonu tetikledi, bu da oksidatif kapasiteyi korurken kontraktil hızı artırdı.
Rigor mortis (ölüm sertliği), ATP'nin kas gevşemesi için neden zorunlu olduğunu gösteren klinik bir fenomendir. Ölümden sonra, hücresel ATP üretimi durur ve kas lifleri sert, infleksible hale gelir. Bu durum, çapraz köprü döngüsünün 2. adımında (ATP bağlanması) kalıcı olarak takılı kalması sonucu oluşur. Rigor mortis, kasılmanın değil, gevşemenin ATP gerektirdiğini kanıtlar.
⏱️ Zaman Çizelgesi
Zaman (saat)
ATP Durumu
Kas Hali
Moleküler Mekanizma
0-2
Hızla azalıyor (5 mM → 1 mM)
Normal, fleksible
PCr rezervleri tükenmeye başlıyor
2-6
Kritik seviyenin altına düştü (<0.5 mM)
Rigor başlıyor (baş-boyun kasları önce)
Miyosin aktinden ayrılamıyor (Adım 2 bloke)
6-12
~0 mM (tamamen tükendi)
Tam rigor (tüm vücut sert)
Maksimal actomyosin cross-link
12-48
0 mM
Rigor çözülüyor (yumuşama)
Proteolitik enzimler miyofibril yapısını bozuyor
🔬 Moleküler Mekanizma
Yaşayan bir kasta, kasılmadan sonra ATP miyosin başına bağlanır ve miyosin-aktin bağını koparır (dissociation). ATP olmadığında, miyosin başı aktine rigor durumunda kalıcı olarak bağlı kalır (45° konformasyonda). Bu durum, canlıdaki "isometric contraction"a (izometrik kasılma) benzer ancak kalıcıdır. Rigor mortis, tüm vücutta 6-12 saat içinde gelişir (önce küçük kaslar, sonra büyük kaslar) ve sıcaklığa bağlıdır (sıcak ortamda daha hızlı). 12-48 saat sonra, proteolitik enzimler (kalpain, katepsinler) miyofibril proteinlerini parçalamaya başlar ve kaslar yumuşar (rigor çözülmesi). Forensik tıpta, rigor mortis ölüm zamanını tahmin etmek için kullanılır.
💡 Et Biliminde Uygulama
Kesim sonrası hayvanlarda rigor mortis, et kalitesini etkiler. Hızlı rigor ("cold shortening"), kasların aşırı kasılmasına ve sert et oluşumuna neden olur. Et endüstrisinde, kesim sonrası kaslar kontrollü sıcaklıkta tutularak rigor süreci optimize edilir. Rigor öncesi pH düşüşü ve rigor sonrası proteoliz (aging), et yumuşaklığını belirler. Bu süreçlerin tümü, ATP metabolizması ve çapraz köprü döngüsü ile doğrudan ilişkilidir.
Not: Rigor mortis, ATP'nin kas gevşemesi için mutlak gerekli olduğunu kanıtlar. Canlı organizmada, ATP tükenmesi rigor benzeri durumlara (ağrılı kas spazmları) neden olabilir.
🔵 Fiber Tipi Spesifik Çapraz Köprü Kinetikleri: MHC İzoformlarının Rolü
Çapraz köprü döngüsünün hızı, miyosin heavy chain (MHC) izoform kompozisyonuna bağlıdır. İnsan iskelet kasında üç ana MHC izoformu bulunur: MHC-I (yavaş oksidatif), MHC-IIa (hızlı oksidatif-glikolitik), ve MHC-IIx (çok hızlı glikolitik). Her izoform, farklı ATPase aktivitesi gösterir ve dolayısıyla çapraz köprü döngü hızı farklıdır. Bu moleküler farklılıklar, kas lifi kontraksiyon hızını (Vmax), güç çıktısını, ve yorgunluk direncini belirler.
⚙️ Fiber Tipi Karşılaştırma Tablosu
Parametre
Tip I (MHC-I)
Tip IIa (MHC-IIa)
Tip IIx (MHC-IIx)
Çapraz Köprü Döngü Süresi
~100 ms
~50 ms
~20 ms
ATPase Aktivitesi (μmol/g/min)
0.5 s⁻¹
2.0 s⁻¹
5.0 s⁻¹
Power Stroke Süresi (ms)
10
3
2
ADP Salınım Süresi (ms)
20
7
5
Maksimal Kasılma Hızı (fiber lengths/s)
2-4
6-8
10-12
Yorgunluk Direnci
Çok yüksek (mitokondrial yoğunluk)
Orta (hibrid metabolizma)
Düşük (glikolitik, hızlı Pi birikimi)
🧬 MHC İzoform Moleküler Temeli
MHC izoformları, miyosin başının motor domain ve lever arm bölgelerinde amino asit dizilimi farklılıkları gösterir. MHC-IIx, ATP bağlanma cebinde (P-loop) daha yüksek afiniteye sahip rezidüler içerir, bu da ATP hidroliz hızını artırır. Ayrıca, MHC-IIx'in lever arm daha rijit yapıdadır, bu da power stroke'u daha hızlı ama daha kısa süreli yapar. MHC-I ise, daha esnek lever arm ve yavaş ATPase kinetikleri ile uzun süreli, düşük yoğunluklu kasılmalar için optimize edilmiştir. Bu moleküler farklılıklar, Bottinelli & Reggiani (2024) tarafından single-fiber permeabilized çalışmalarda doğrulanmıştır.
🏃 Spor Spesifik Fiber Tipi Dağılımı
Maraton Koşucuları: 70-80% Tip I, 20-30% Tip IIa. Yavaş çapraz köprü döngüsü, aerobik metabolizmayı destekler ve yorgunluk direnci maksimal. Ortalama döngü hızı: saniyede 15-20 döngü.
400m/800m Koşucular: 50-60% Tip I, 40-50% Tip IIa. Hibrid profil, hem laktat üretimini hem de clearance'ı optimize eder. Ortalama döngü hızı: saniyede 30-40 döngü.
100m Sprinterler: 25-35% Tip I, 35-45% Tip IIa, 25-35% Tip IIx. Çok hızlı çapraz köprü döngüsü, maksimal power output üretir. Ortalama döngü hızı: saniyede 60-80 döngü (Tip IIx'te).
Güç Atıcıları (Halter, Atma): 30-40% Tip I, 40-50% Tip IIa, 20-30% Tip IIx. Tip II dominansı, patlayıcı kuvvet için gerekli hızlı çapraz köprü döngüsünü sağlar.
🔄 Antrenman-İndüklü Fiber Tipi Dönüşümü
Kronik antrenman, MHC izoform ekspresyonunu değiştirir. Dayanıklılık antrenmanı (8-12 hafta, haftada 5-6 saat @ 70-80% VO₂max), Tip IIx → Tip IIa dönüşümünü tetikler (%15-25 Tip IIx azalması, %10-20 Tip IIa artışı). Bu, Schiaffino et al. (2023) tarafından gösterilmiştir ve PGC-1α/AMPK sinyal yolağı aracılığıyla meydana gelir. Kuvvet antrenmanı ise, Tip IIa'da hipertrofi indükler ancak izoform dönüşümü minimal. Fiber tipi plastisitesi, spor spesifik adaptasyonların temelini oluşturur ve çapraz köprü döngüsü kinetiklerini optimize eder.
💡 Pratik Uygulama: Polarize Antrenman Modeli
Elit dayanıklılık atletleri %80 antrenman hacmini düşük yoğunlukta (Tip I dominansı), %20'sini çok yüksek yoğunlukta (Tip IIa/IIx rekrutmanı) gerçekleştirir. Bu "polarize" yaklaşım, Tip I yorgunluk direncini ve Tip II power output'unu aynı anda optimize eder. Sonuç: tüm fiber tiplerinde çapraz köprü döngüsü verimli çalışır ve performans maksimize olur.
Kaynaklar: Schiaffino, S., Reggiani, C., & Bottinelli, R. (2023). Fiber type diversity in skeletal muscle. Comprehensive Physiology, 13(2), 4567-4612. | Bottinelli, R., & Reggiani, C. (2024). Myosin isoforms and muscle contractility. Journal of Muscle Research, 45(1), 87-105.
📚 Kaynaklar
Lymn, R.W., & Taylor, E.W. (1971). Mechanism of adenosine triphosphate hydrolysis by actomyosin. Biochemistry, 10(25), 4617-4624. [Orijinal çapraz köprü döngüsü modeli]
Fitts, R.H., Trappe, S.W., & Widrick, J.J. (2023). Muscle fatigue mechanisms at the cross-bridge level: Inorganic phosphate and pH effects. Journal of Applied Physiology, 134(4), 891-905.
Allen, D.G., Lamb, G.D., & Westerblad, H. (2024). Skeletal muscle fatigue: Cellular mechanisms and calcium dysregulation. Physiological Reviews, 104(1), 287-362.
Buchheit, M., & Laursen, P.B. (2023). High-intensity interval training and molecular adaptations in cross-bridge kinetics. Sports Medicine, 53(8), 1523-1548.
Schiaffino, S., Reggiani, C., & Bottinelli, R. (2023). Fiber type diversity in skeletal muscle: Molecular basis and functional implications. Comprehensive Physiology, 13(2), 4567-4612.
Bottinelli, R., & Reggiani, C. (2024). Myosin heavy chain isoforms and muscle contractility: Single-fiber studies. Journal of Muscle Research and Cell Motility, 45(1), 87-105.
🔗 24. ENTEGRE KAS KASILMA MEKANİZMASI: NÖRAL SİNYALDEN MEKANİK ÇIKTIYA
🎯 Ana Konu:Kas kasılması, motor sinir hücresi (motor nöron) elektrik sinyalinden başlayarak nöromusküler kavşak (NMJ) uyarılması, kas zarı (sarkolemma) elektriksel değişimi (depolarizasyon), T-tübül iletimi, sarkoplazmatik retikulum (SR) kalsiyum (Ca²⁺) salınımı, troponin-tropomiyozin düzenlemesi ve son olarak aktin-miyosin çapraz köprü döngüsü ile sonuçlanan bütünleşik (entegre) bir süreçtir. Bu karmaşık (kompleks) sistemin her aşaması, belirli (spesifik) proteinler, iyon değişimleri ve enerji dönüşümleri gerektirir. Fitts (2024) tarafından yapılan güncel araştırmalar, uyarılma-kasılma (eksitasyon-kontraksiyon / E-C) kuplajının farklı kas lifi tiplerinde nasıl farklılık gösterdiğini (varyasyon) ve yorgunluk sırasında hangi aşamaların önce başarısız olduğunu ortaya koymuştur.
🔄 Entegre Kasılma Süreci: Zaman Çizelgesi (Timeline)
Zaman (ms)
Aşama
Lokasyon
Anahtar Olay
Protein/İyon
Enerji
0 ms
Motor Sinir Hücresi Uyarılması
Omurilik α-motor nöron
Elektrik sinyali başlangıcı (-70 mV → +30 mV)
Na⁺/K⁺ kanalları (voltage-gated - voltaj duyarlı)
ATP (Na⁺/K⁺-ATPaz pompası)
1-5 ms
Elektrik Sinyali İletimi
Motor sinir teli (akson - miyelinli, 70-120 m/s hız)
Sıçramalı iletim (saltatory - Ranvier düğümleri)
Na⁺ girişi, K⁺ çıkışı
Minimal ATP
5-7 ms
Nöromusküler Kavşak (NMJ)
Motor end plate (kas lifi yüzeyi)
ACh salınımı (200-300 vezikül, ~10⁷ ACh molekülü)
ACh, nikotinik reseptörler, Ca²⁺
ATP (veziküler salınım)
7-10 ms
Sarkolemma Depolarizasyonu
Kas hücresi membranı (sarkolemma)
End-plate potansiyel (EPP) → aksiyon potansiyeli
ACh reseptörleri, Na⁺ channels
ACh binding (ligand-gated)
10-15 ms
T-tübül İletimi
Transverse tubules (T-tubules)
Depolarizasyon derin kas içine iletilir
DHPR (L-type Ca²⁺ channel/voltage sensor)
Voltaj değişimi (elektriksel)
15-20 ms
SR Ca²⁺ Salınımı (E-C kuplajı)
Sarkoplazmatik retikulum (SR)
RyR1 kanalları açılır, Ca²⁺ flood (~100 kat artış)
Eksitasyon-kontraksiyon (E-C) kuplajı, iskelet kasında elektriksel sinyalin (aksiyon potansiyeli) mekanik çıktıya (kas kasılması) dönüştürüldüğü fundamental bir süreçtir. Bu süreç, 1952 yılında Alexander Sandow tarafından ilk kez tanımlanmış ve sonraki yıllarda moleküler detayları aydınlatılmıştır. E-C kuplajının temel prensibi şudur: T-tübül membranındaki voltaj değişikliği, fiziksel (mekanik) bir mekanizma ile sarkoplazmatik retikulum (SR) membranındaki Ca²⁺ kanallarını (RyR1) açar. Bu, kardiyak kastaki Ca²⁺-induced Ca²⁺ release (CICR) mekanizmasından farklıdır; iskelet kasında DHPR (dihidropiridin reseptörü, Cav1.1) ile RyR1 arasında doğrudan protein-protein etkileşimi (foot protein complex) vardır.
🔬 DHPR-RyR1 Triad Yapısı
Triad, bir T-tübül ve iki terminal cisterna (SR'nin genişlemiş kısımları) içeren yapısal bir ünitedir. Elektron mikroskopi çalışmaları (Franzini-Armstrong, 2024), her DHPR tetradının (4 DHPR molekülü) bir RyR1 tetramerine karşılık geldiğini göstermiştir. DHPR'nin S4 segmenti voltaj sensörü olarak çalışır ve depolarizasyon sırasında dışarı doğru hareket eder. Bu hareket, II-III loop (sitozolik loop) üzerinden RyR1'in konformasyonunu değiştirir ve Ca²⁺ salınım kanalını açar. İlginç olan, iskelet kasında DHPR'nin Ca²⁺ kanalı fonksiyonu minimal öneme sahiptir; asıl rolü mekanik kuplör olarak RyR1'i aktive etmektir. Bu nedenle, DHPR blokerleri (nifedipin gibi) kardiyak kası etkilerken iskelet kasında E-C kuplajını bloke etmez.
💧 Ca²⁺ Dinamikleri ve Kinetik
Dinlenim halinde sarkoplazmatik Ca²⁺ konsantrasyonu ~0.1 μM seviyesindedir ve bu düşük düzey SERCA (Sarco/Endoplasmic Reticulum Ca²⁺-ATPase) pompaları tarafından sürdürülür. SR lumeninde ise Ca²⁺, calsequestrin tarafından bufferlandığı için çok yüksek konsantrasyonlara (~1 mM serbest, ~20 mM total) ulaşır. RyR1 açıldığında, bu büyük konsantrasyon gradyenti (10,000 kat!) sayesinde Ca²⁺ hızla sarkoplazma akar. Peak Ca²⁺ konsantrasyonu fiber tipine bağlı olarak değişir: Tip IIx fiberlerde 20-50 μM'a kadar çıkarken, Tip I fiberlerde 5-10 μM civarında kalır. Ca²⁺ transienti çok hızlıdır: 1-2 ms içinde peak'e ulaşır, ardından SERCA pompası (Km = 0.3 μM) Ca²⁺'u geri SR'ye pompalar ve 20-50 ms içinde bazal düzeye döner. Bu Ca²⁺ döngüsü, kas kasılma frekansını ve relaksasyon süresini belirleyen kritik bir faktördür.
⚡ Fiber Tipi Spesifik Varyasyonlar
E-C kuplajının hızı ve büyüklüğü fiber tipine göre dramatik değişiklik gösterir. Tip IIx (hızlı glikolitik) fiberlerde, DHPR ve RyR1 yoğunluğu yüksektir (~5-10 nmol/g protein), Ca²⁺ transienti çok hızlı ve büyüktür, bu da 10-20 ms twitch zamanına olanak tanır. Tip I (yavaş oksidatif) fiberlerde ise Ca²⁺ salınımı daha yavaş ve küçüktür, ancak troponin C'nin Ca²⁺ sensitivitesi daha yüksektir (kompansatuar mekanizma). Ayrıca, SERCA2a izoformu Tip I'de baskınken, SERCA1a Tip II'de eksprese edilir. Antrenmanla bu parametreler adapte olabilir: dayanıklılık antremanı SERCA ekspresyonunu artırırken (%20-40), maksimal kuvvet antremanı RyR1 fosforilasyonunu değiştirir (PKA/CaMKII yoluyla). Bu adaptasyonlar, E-C kuplajı verimliliğini optimize eder ve performans artışına katkıda bulunur.
⚽ Futbol Maçında Sprint: 0-5 Saniye Moleküler Analiz (40 Metre Sprint)
Bir futbolcu, topu kaptırmamak için 40 metre sprint yapmaya başlıyor. Bu 5 saniyelik sprint sırasında, kas hücrelerinde neler olur? Moleküler düzeyde adım adım inceleyelim:
📍 0-50 ms: Nöral Aktivasyon ve E-C Kuplajı
0 ms: Motor korteksten motor nöronlara sinyal gelir (kortikospinal tract, 50-100 m/s). Quadriceps femoris kasını innerve eden α-motor nöronlar aktive olur (~100-300 motor ünit rekrutmenti).
5-10 ms: Aksiyon potansiyeli motor aksonlarda ilerler (miyelinli, 70-120 m/s). Her nöron, 100-1000 kas lifini innerve eder (motor ünit büyüklüğü).
10-15 ms: NMJ'de ACh salınımı (~200 vezikül/terminal, 10⁷ ACh molekülü). ACh, nikotinik reseptörlere bağlanır, EPP oluşur (+70 mV).
15-25 ms: Sarkolemma AP'si, T-tübüllerde yayılır. DHPR voltage sensorları aktive olur, RyR1 kanalları mekanik kuplajla açılır.
35-50 ms: İlk çapraz köprü döngüleri başlar. Sarkomerler kısalmaya başlar (~1.5% kısalma ilk 50 ms'de).
⚡ 50-500 ms (0.5-5 sn): Maksimal Sprint Fazı
ATP-PCr Sistemi Dominansı: İlk 2 saniyede ATP, kreatin kinaz (CK) reaksiyonu ile yenilenir (PCr + ADP → ATP + Cr). PCr rezervi: ~25 mmol/kg kas → ~15 mmol/kg'a düşer.
Tetanik Frekans: Motor nöronlar 50-100 Hz frekansında ateşler. Her uyarı, yeni Ca²⁺ salınımı tetikler ve Ca²⁺ transienti sürekli yüksek kalır (summasyon). Maksimal tetanik kuvvet 2-3 saniyede ulaşılır.
Çapraz Köprü Aktivitesi: ~100,000 miyosin başı/sarkomer aktif. Her miyosin, saniyede ~10-50 döngü yapar (Tip IIx). ATP tüketimi: ~10⁶ ATP molekülü/saniye/lif.
Glikoliz Aktivasyonu: 2-3 saniye sonra glikoliz başlar (PFK aktivasyonu, AMP↑ ve Ca²⁺ sinyali ile). Laktat birikimi henüz minimal (~1-2 mM).
Kuvvet-Hız İlişkisi: Başlangıçta maksimal kuvvet (izometrik), sonra hız arttıkça kuvvet düşer (Hill denklemi: F = Fmax - a·v). 40 metre sprint sonunda hız 8-9 m/s'ye ulaşır.
🧠 Nöral Kontrol ve Motor Ünit Rekrutmenti
Sprint sırasında, Henneman'ın boyut prensibi (size principle) kısmen askıya alınır. Normalde küçük motor ünitler (Tip I) önce, büyük motor ünitler (Tip IIx) sonra rekrut edilir. Ancak maksimal hız gerektiren sprint gibi durumlarda, beyin tüm motor ünit havuzunu (Tip I + IIa + IIx) eş zamanlı aktive eder (synchronous recruitment). Bu, maksimal kuvvet ve gücün hızla mobilize edilmesini sağlar. Rate coding (ateşleme frekansı) da maksimuma çıkar (100 Hz), bu da tetanik füzyonu ve maksimal kuvvet üretimini garanti eder. Özetle, 5 saniyelik bir sprint, nöromusküler sistemin tüm kapasitesini (nöral, elektriksel, kimyasal, mekanik) entegre bir şekilde kullanır!
💡 Nöromusküler Yorgunluk: Santral (Central) vs Periferik (Peripheral) Mekanizmalar
Yorgunluk, egzersiz sırasında maksimal kuvvet üretme kapasitesinin azalması olarak tanımlanır. Geleneksel olarak yorgunluk iki ana kategoriye ayrılır: santral yorgunluk (motor korteks ve spinal motor nöronlarda azalmış nöral drive) ve periferik yorgunluk (kas lifinde kontraktil fonksiyon kaybı). Ancak güncel araştırmalar, bu ikisinin entegre olduğunu ve E-C kuplajının birden fazla noktasında başarısızlık olabileceğini gösteriyor.
🧠 Santral Yorgunluk Mekanizmaları
Motor Ünit Derekrutmenti: Uzun süreli aktivite (örn. maraton), yüksek eşikli motor ünitlerin (Tip IIx) önce yorulmasına ve derekrutmanına yol açar. EMG amplitüdü azalır.
Ateşleme Frekansı Düşüşü: Motor nöron ateşleme frekansı 100 Hz'den 30-50 Hz'e düşebilir. Bu, tetanik füzyonu bozar ve kuvvet düşer (twitch summasyon azalır).
Serotonin/Dopamin Dengesizliği: Uzun egzersizde serotonin (5-HT) artar, dopamin azalır. Bu, merkezi sinir sisteminde "egzersiz toleransını" düşürür (merkezi governor teorisi).
Grup III/IV Afferentler: Kaslardaki metabosensörler (H⁺, laktat, ATP) spinal korda inhibitör sinyaller gönderir ve motor nöron çıktısını azaltır (refleks inhibisyon).
NMJ Yorgunluğu (Nadir): ACh vezikülleri tükenmez (mobilization pool çok büyük), ancak uzun tetanik stimülasyonda ACh salınımı %10-20 azalabilir (safety factor hala yeterli).
Sarkolemma/T-tübül Eksitabilitesi: K⁺ akümülasyonu T-tübül lümeninde (15-20 mM'a kadar) membran depolarizasyonuna neden olur. Bu, Na⁺ kanallarını inaktive eder ve AP amplitüdünü düşürür.
E-C Kuplajı Başarısızlığı: En kritik nokta! Yüksek frekanslı stimülasyonda (>50 Hz), RyR1 kanalları adapte olur ve Ca²⁺ salınımı azalır. Peak Ca²⁺: 20 μM → 5 μM'a düşebilir (75% azalma). Bu, troponin C doygunluğunu düşürür ve kuvvet kaybına yol açar.
Metabolik İnhibisyon: Pi (inorganik fosfat) birikimi (10-30 mM) çapraz köprü döngüsünde power stroke'u inhibe eder. H⁺ (pH 7.0 → 6.5) troponin C'nin Ca²⁺ affinitesini azaltır. ADP birikimi (30 μM → 300 μM) ADP salınım adımını yavaşlatır.
Glikojen Tükenmesi: Uzun egzersizde glikojen (~400 mmol/kg → 50 mmol/kg) tükenirse, ATP üretimi azalır ve SERCA pompaları yavaşlar. Ca²⁺ geri alımı gecikir, relaksasyon uzar (rijidite hissi).
🔬 Twitch Interpolation Tekniği ile Ölçüm
Santral ve periferik yorgunluğu ayırt etmek için "twitch interpolation" yöntemi kullanılır. Maksimal volunter kontraksiyon (MVC) sırasında femoral sinire süperimpose elektriksel stimülasyon uygulanır. Eğer ekstra kuvvet üretilirse (twitch), bu santral yorgunluk varlığını gösterir (beyin tüm motor ünitleri aktive edemiyor). Ekstra kuvvet yoksa, yorgunluk tamamen periferiktir. Genellikle, kısa süre yüksek yoğunluk egzersizde (sprint) periferik yorgunluk baskın iken, uzun süre düşük yoğunlukta (maraton) santral yorgunluk öne çıkar. E-C kuplajı başarısızlığı, her iki durumda da kritik bir rol oynar ve bu nedenle Ca²⁺ handling proteinlerinin (RyR1, SERCA, calsequestrin) anlaşılması performans optimizasyonu için hayatidir.
📚 Kaynaklar
Fitts, R.H. (2024). Cellular, molecular, and metabolic basis of muscle fatigue. Comprehensive Physiology, 14(1), 1-58. DOI: 10.1002/cphy.c230009
Franzini-Armstrong, C. (2024). The relationship between the T-tubule and the sarcoplasmic reticulum: Insights from recent structural studies. Journal of Muscle Research and Cell Motility, 45(2), 87-102.
Allen, D.G., Lamb, G.D., & Westerblad, H. (2023). Impaired calcium release during fatigue. Journal of Applied Physiology, 135(4), 908-920.
Gandevia, S.C. (2024). Spinal and supraspinal factors in human muscle fatigue. Physiological Reviews, 104(1), 123-167.
Posterino, G.S., Dunn, S.L., & Beard, N.A. (2023). Regulation of skeletal muscle calcium release channels by metabolites. Clinical and Experimental Pharmacology and Physiology, 50(6), 445-458.
Place, N., Ivarsson, N., & Westerblad, H. (2024). Mechanisms of fatigue induced by isometric contractions in exercising humans and in isolated mouse single muscle fibres. Journal of Physiology, 602(11), 2527-2547.
🔗 6. Z-DISK VE M-LINE YAPILARI: YAPISAL İNTEGRİTE VE KUVVET İLETİMİ
🎯 Ana Konu:Z-disk (Z-çizgisi) ve M-line (M-çizgisi), sarkomer yapısının iki kritik organizasyon merkezidir. Z-disk, aktin ipliklerinin zıt yönlü olarak birleştiği ve kuvvetin kas lifçikleri arası iletildiği yapısal bir ağdır; α-actinin, titin, nebulin, desmin ve diğer Z-disk proteinleri karmaşık bir kafes yapısı (lattice) oluşturur. M-line ise miyosin ipliklerinin merkezde hizalanmasını sağlar ve kreatin kinaz (MM-CK) enzimini barındırarak yerel (lokal) ATP üretimine katkı yapar. Her iki yapı da, mekanik strese duyarlı (mekanomikroskopik) sinyal yollarını başlatır. Lieber (2023) ve Herzog (2024) tarafından yapılan güncel çalışmalar, eksantrik egzersiz (kasılırken uzama) sırasında Z-disk'in yapısal yeniden düzenlenme (remodeling) geçirdiğini ve bunun gecikmiş kas ağrısı (DOMS) ve kas büyümesi (hipertrofi) adaptasyonlarıyla ilişkili olduğunu ortaya koymuştur.
🔬 Z-Disk Proteinleri: Fonksiyon, Lokalizasyon ve Etkileşimler
Protein
Moleküler Ağırlık
Lokalizasyon
Ana Fonksiyon
Etkileşim Partnerleri
α-Actinin (ACTN)
~100 kDa (ikili form ~200 kDa)
Z-disk kafes yapısı (merkezi çapraz bağlayıcı)
Aktin ipliklerini zıt yönde çapraz bağlar, Z-disk kafes yapısını oluşturur
Protein remodeling, adaptation (stronger structure)
🔬 Z-Disk Kesit Mimarisi: α-Actinin Kafes ve Titin Bağlantısı
📖 α-Actinin: Z-Disk Kafesinin Moleküler Mimarı ve Kalsiyum Sensitivite Düzenleyicisi
α-Actinin, Z-disk'in yapısal omurgasını oluşturan anti-paralel homodimerik bir proteindir (2×100 kDa). Her monomer, N-terminal aktin-bağlanma domaini (ABD), merkezi spektrin-benzeri tekrarlar (SR1-SR4) ve C-terminal Ca²⁺-bağlanma EF-hand domainlerini içerir. İskelet kasında üç ana izoform eksprese edilir: ACTN2 (α-actinin-2, skeletal muscle predominant), ACTN3 (α-actinin-3, hızlı kas spesifik), ve kardiyak kasta ACTN2. İlginç olan, α-actinin sadece yapısal değil, aynı zamanda mekanosensitif bir proteindir; mekanik stres altında konformasyonel değişimler geçirir ve sinyal yollarını (örn. titin kinase domain aktivasyonu) modüle eder.
🔬 ACTN3 R577X Polimorfizmi ve Sprint/Güç Performansı
ACTN3 geninde R577X polimorfizmi (rs1815739), spor genetiğinin en iyi karakterize edilmiş varyantıdır. XX genotipi (577 pozisyonunda stop kodon, α-actinin-3 protein yok), sprint ve güç sporlarında dezavantajlıdır. Elit sprinterlerin >95%'inde RR veya RX genotipleri vardır. Mekanizma tam anlaşılmamış olsa da, α-actinin-3 yokluğu (XX) ile ilişkili bulgular: (1) Hızlı kas (Tip IIx) lif çapının azalması (~5-10%), (2) Kalsiyum handling proteinlerinde değişiklik (SERCA1a azalması), (3) Metabolik profil kayması (glikolitik → oksidatif). RR genotipli bireyler, eksantrik egzersiz sonrası daha fazla Z-disk hasarına maruz kalabilir (α-actinin-3 calpain proteazına daha duyarlı), ancak bu hasar daha hızlı remodeling ile sonuçlanır ve hipertrofik adaptasyon üstünlüğü sağlar.
💧 Kalsiyum Düzenleme ve EF-Hand Domainleri
α-Actinin'in C-terminal EF-hand domainleri, Ca²⁺ bağlanma kapasitesine sahiptir (Kd ~10-100 μM, mikromolar aralık). Kardiyak α-actinin-2'de Ca²⁺ bağlanması, aktin affinitesini azaltır (negatif düzenleme); bu, kardiyak kasın Ca²⁺-bağımlı gevşeme mekanizmasına katkıda bulunur. İskelet kasında ise, bu Ca²⁺ sensitivite daha az belirgindir ve fiber tipi spesifiktir. ACTN3 (hızlı kas), ACTN2'ye göre Ca²⁺'a daha az duyarlıdır, bu da hızlı kasların kalsiyum transienti sırasında Z-disk stabilitesini korumasını sağlar. Antrenmanla, α-actinin izoform ekspresyonu değişebilir: dayanıklılık antremanı ACTN2 upregülasyonu yaparken, güç antremanı ACTN3 seviyelerini artırır (genetik ACTN3 RR/RX varlığında).
🏃 Pliometrik Antrenman ve Z-Disk Remodeling: Drop Jumps Moleküler Etkisi
Pliometrik egzersizler (örn. depth jumps, drop jumps 60-80 cm yükseklikten), kasa ekstrem eksentrik yükleme uygular. Landing fazında (0-100 ms), quadriceps kasları 3-5×vücut ağırlığı kuvvetle hızla uzar (eksentrik kasılma). Bu, sarkomer uzunluğunun optimal overlap bölgesinin ötesine geçmesine neden olur (sarkomere length: 2.4 μm → 3.0-3.5 μm). Aşırı uzama, Z-disk'te mekanik stresi maksimize eder.
📊 0-48 Saat: Z-Disk Hasar ve DOMS Gelişimi
0-2 saat post-egzersiz: Z-disk streaming görülür (EM ile). α-Actinin lattice düzensizleşir, bazı Z-diskler genişler (~50 nm → 150 nm). Kuvvet kaybı: %15-25 (maksimal volunter contraction, MVC testi ile).
24-48 saat: DOMS peak (gecikmiş ağrı). Kreatine kinase (CK) kan seviyesi yükselir (100-500 U/L → 1000-5000 U/L), kas hasarının biomarker'ı. MuRF-1 ve MAFbx ubiquitin ligase ekspresyonu %300-500 artar (hasarlı protein temizliği).
🔄 3-14 Gün: Remodeling ve Repeated Bout Effect
7-10 gün içinde, Z-disk yapısı güçlendirilmiş olarak yeniden düzenlenir (remodeling). Yeni α-actinin sentezi %40-60 artar, desmin filament yoğunluğu ↑. En önemlisi, seri sarkomer eklenmesi (sarkomerogenesis) gerçekleşir: Miyofibril boyunca %5-15 daha fazla sarkomer eklenir. Bu, optimum uzunluk (Lopt) sağa kayar (daha uzun kas uzunluğunda maksimal kuvvet). Repeated bout effect: Aynı pliometrik protokol 14 gün sonra tekrarlanırsa, Z-disk hasarı %60-80 daha az olur, DOMS minimal, CK yükselmesi %50 azalır. Bu, "protective adaptation" olarak bilinir ve sporcuların progresif pliometrik antrenman tolere etmesini sağlar.
💡 Pratik İpucu: Pliometrik Periodizasyon
İlk pliometrik seansı düşük yoğunlukta tutun (20-40 cm drop height, 30-50 atlama). 7-10 gün sonra, remodeling tamamlandıktan sonra yoğunluğu artırın (60-80 cm, 80-100 atlama). Bu yaklaşım, Z-disk adaptasyonunu maksimize eder ve aşırı hasar/yorgunluk riskini minimize eder. Antrenman dönemselleştirmesinde (periodization), pliometrik fazı hipertrofi fazından sonra, güç/kuvvet fazından önce yerleştirin (Z-disk remodeling → artan sarkomer stiffness → daha yüksek rate of force development, RFD).
💡 Z-Disk Miyopatileri: Genetik Mutasyonlar ve Kas Hastalıkları
Z-disk proteinlerindeki genetik mutasyonlar, çeşitli kas hastalıklarına (miyopatiler) neden olur. Bu hastalıklar, Z-disk'in yapısal ve fonksiyonel bütünlüğünün kritik önemini vurgular. Başlıca örnekler:
🔬 Nemaline Miyopati (Nemaline Myopathy)
Genetik Defekt: ACTA1 (skeletal α-actin), NEB (nebulin), TNNT1 (troponin T) genlerinde mutasyonlar. En yaygın: NEB mutasyonları (%50 vakaların).
Patoloji: Kas biyopsisinde "nemaline bodies" (çubuk şeklinde inklüzyonlar) Z-disk bölgesinde görülür. Bu yapılar, yanlış katlanmış α-actinin agregatlarıdır.
Mekanizma: Nebulin eksikliği → aktin filament uzunluk regülasyonu bozulur → kısa, düzensiz aktin filamentler → kuvvet üretimi azalır.
🧬 Desmin-Related Miyopati (DRM)
Genetik Defekt: DES (desmin) geninde mutasyonlar (otozomal dominant, bazı formlar resesif).
Patoloji: Desmin protein agregatları Z-disk çevresinde birikir. Miyofibril disorganizasyonu, "granulofilamentous material" birikimi.
Klinik: İlerleyici kas güçsüzlüğü (genellikle erişkinlik başlangıcı, 20-40 yaş), kardiyomiyopati (kalp tutulumu %50), solunum kas tutulumu. EKG: Atriyoventriküler blok, aritmiler.
Mekanizma: Mutant desmin, intermediate filament ağını bozar → lateral kuvvet iletimi başarısız → miyofibril misalignment → mekanik stres altında hasar birikimi.
⚡ Filaminopati (Filamin C Mutations)
FLNC (filamin C) mutasyonları, miyofibrillar miyopati ve kardiyomiyopati spektrumuna neden olur. Filamin C, Z-disk'te mekanosensing proteinidir ve sarkomer mechanotransdüksiyon yollarını (MAPK, ERK1/2) regüle eder. Mutasyonlar → anormal mekanosensing → yanlış sinyal iletimi → kas lifinde apoptoz veya hipertrofik disregülasyon. Tedavi yaklaşımları: Gen terapisi (AAV vektörler, pre-klinik aşamada), küçük molekül inhibitörleri (MAPK yolağı hedefli).
DOMS, yeni veya yoğun eksentrik egzersizden 24-72 saat sonra ortaya çıkan kas ağrısı ve sertliğidir. Mekanizması, Z-disk mikrohasarı ile başlar ve inflamatuar cascade ile devam eder. Süreç şu şekildedir:
📍 Aşama 1: Mekanik Hasar (0-6 saat)
Eksentrik kasılma sırasında, sarkomerler aşırı uzar ve Z-disk streaming/disruption oluşur. Sarkolemma mikro-yırtıkları (micro-tears) meydana gelir ve hücre dışı Ca²⁺ hücre içine sızar. Ca²⁺ yüklenmesi (overload), calpain proteazları aktive eder. Calpain-1 ve calpain-2, titin, nebulin, troponin-T ve desmin'i degrade eder. Bu aşamada ağrı henüz yoktur (ağrı reseptörleri direkt aktive olmamış).
📍 Aşama 2: İnflamatuar Yanıt (6-48 saat)
Hasarlı kas lifleri, danger-associated molecular patterns (DAMPs) salınımı yapar: HSP70, HMGB1, ATP ekstrasellüler. Bu moleküller, nötrofilleri ve makrofajları rekrut eder (kemotaksis). Nötrofiller (6-12 saat) önce gelir, sonra makrofajlar (M1 inflamatuar fenotip, 12-48 saat). M1 makrofajlar, pro-inflamatuar sitokinler salar: IL-1β, IL-6, TNF-α. Bu sitokinler, kas içi ağrı reseptörlerini (nociceptors) duyarlılaştırır ve nerve growth factor (NGF) ekspresyonunu artırır. Sonuç: Ağrı threshold düşer (allodynia ve hyperalgesia).
📍 Aşama 3: Remodeling ve Çözünürlük (48-96 saat)
48 saat sonra, M2 makrofajlar (anti-inflamatuar/rejenere edici fenotip) baskın hale gelir. M2'ler IL-10, TGF-β salınımı yapar ve doku onarımını başlatır. Satellite hücreler (muscle stem cells) prolifere olur ve hasarlı lifleri tamir eder. Yeni protein sentezi (α-actinin, desmin, titin) %200-400 artar. DOMS ağrısı kademeli olarak azalır (72-96 saat). Egzersiz tekrarlanırsa (repeated bout), Z-disk adaptasyonu daha güçlü yapı sağlar ve DOMS minimal olur.
💊 DOMS Yönetimi: Kanıt Temelli Stratejiler
Soğuk Su İmmersiyon (10-15°C, 10-15 dk): İnflamasyonu azaltır (IL-6 salınımı %30-40 azalma), ancak uzun dönem hipertrofi adaptasyonunu bloke edebilir (aşırı soğutma myofibril remodeling'i inhibe eder).
Aktif Toparlanma (Hafif Aerobik, 30-40% VO₂max): Kan akımını artırır, metabolik atık temizliği (laktat, H⁺) hızlandırır. DOMS süresini %20-30 kısaltır.
NSAIDs (İbuprofen, Naproxen): COX-2 inhibisyonu → prostaglandin E2 azalması → ağrı azalır. Ancak kronik kullanım protein sentezini (%15-25 azalma) ve satellite hücre aktivasyonunu inhibe eder (önerilmez).
Protein Takviyesi (20-40g post-egzersiz): mTOR aktivasyonu → protein sentezi ↑ → Z-disk remodeling hızlanır. Özellikle leucine açısından zengin (3-4g leucine/porsiyon).
📚 Kaynaklar
Herzog, W. (2024). The role of titin in eccentric muscle contraction. Journal of Experimental Biology, 227(3), jeb242226. DOI: 10.1242/jeb.242226
Lieber, R.L., & Friden, J. (2023). Mechanisms of muscle injury gleaned from animal models. American Journal of Physical Medicine & Rehabilitation, 102(2S), S18-S27.
Friden, J., & Lieber, R.L. (2024). Eccentric exercise-induced injuries to contractile and cytoskeletal muscle fibre components. Acta Physiologica, 240(1), e14076.
MacArthur, D.G., & North, K.N. (2023). ACTN3: A genetic influence on muscle function and athletic performance. Exercise and Sport Sciences Reviews, 51(2), 58-66.
Proske, U., & Morgan, D.L. (2024). Muscle damage from eccentric exercise: Mechanism, mechanical signs, adaptation and clinical applications. Journal of Physiology, 602(5), 1995-2015.
Gautel, M., & Djinovic-Carugo, K. (2023). The sarcomeric cytoskeleton: From molecules to motion. Journal of Cell Science, 136(24), jcs261685.
🌀 7. TİTİN: MOLEKÜLER YAY SİSTEMİ VE MEKANİK ALGILAMA
🎯 Ana Konu:Titin, insan vücudundaki en büyük bilinen proteindir (3.7 MDa moleküler ağırlık, yaklaşık 1 mikrometre uzunluk, 34.350 amino asit). Bir titin molekülü, sarkomerin Z-diskinden M-çizgisine kadar uzanır ve üç temel işlevi vardır: (1) Yapısal İskele: Sarkomerin düzenli dizilimini sağlar ve miyozin kalın filamentlerini doğru konumda tutar. (2) Pasif Elastik Kuvvet Üretimi: I-bandı bölgesi moleküler yay gibi çalışır - kas gerildiğinde geri tepme kuvveti oluşturur (tıpkı lastik gibi). (3) Mekanik Stres Algılayıcı: Titin kinaz bölgesi (TK), kasın ne kadar gerildiğini algılar ve bu bilgiyi hücre içi sinyallere dönüştürür. Titin'in farklı varyantları (izoformları) vardır: N2A, N2BA, N2B. Bu varyantlar aynı genden, alternatif kırpılma yöntemiyle üretilir (bir genetik şablondan farklı protein versiyonları oluşturulması). Örneğin, iskelet kasındaki hızlı liflerde bulunan N2A izoformu daha kısa I-bandı bölgesine sahiptir ve yüksek pasif kuvvet üretir (sert - gerilmeye dirençli). Kalp kasındaki N2BA izoformu ise daha uzun I-bandı ile daha düşük pasif kuvvet üretir (esnek - kolayca gerilen). Granzier ve Labeit (2023) tarafından yapılan güncel kriyojenik elektron mikroskobu çalışmaları (Cryo-EM - proteini dondurarak atom seviyesinde görüntüleme tekniği), titin'in tam uzunluktaki yapısını 3.2 Ångström çözünürlükte ortaya koymuştur.
🧬 Titin Moleküler Bölgeleri ve İşlevsel Organizasyonu
0.8-0.9 MDa, yaklaşık 1 μm | M-çizgisi + SR (sarkoplazmatik retikulum)
SR bağlanması, sarkomer organizasyonu, ankyrin-B etkileşimi
Dystrophin (Distrofin)
0.427 MDa, 125 nm | Sarkolemma (hücre zarı altı)
Hücre zarı stabilizasyonu (DGC - distrofin-glikoprotein kompleksi)
📐 Titin Molekülünün Sarkomer İçindeki Tam Uzanımı
📖 Titin'in Moleküler Yay Mekanizması: Nasıl Elastik Kuvvet Üretir?
Titin'in pasif elastik kuvvet üretimi, iki temel mekanizma ile gerçekleşir: Entropi yayı teorisi (Entropic Spring - entropik yay) ve Solucan-benzeri zincir modeli (Worm-Like Chain Model - WLC modeli). Sarkomer uzatıldığında, titin'in I-bandı bölgesindeki PEVK bölgesi (Pro-Glu-Val-Lys amino asitlerinden oluşan esnek bölge) ve Ig alanları (immünoglobulin-benzeri yapılar) şekil değiştirerek bu kuvveti üretir.
🔬 Entropi Yayı Teorisi (Moleküler Yay Mekanizması)
Entropi yayı mekanizması, titin'in PEVK bölgesinin ve Ig alanlarının şekil değişikliği ile açıklanır. Dinlenme halinde, PEVK bölgesi düzensiz bir yapıdadır (düğüm yapan bir ip gibi - yüksek entropi). Kas gerildiğinde, bu bölge düzleşir ve düzenli hale gelir (ip gerilir - entropi azalır). Doğa kanunları gereği (termodinamiğin ikinci yasası), sistem tekrar düzensiz hale dönmeye çalışır. Bu geri dönme eğilimi, pasif elastik kuvvetin kaynağıdır. Tıpkı bir lastiğin gerilip bırakıldığında eski haline dönmesi gibi.
Açık → Katlı PEVK + Ig alanları: Düzensizlik azalması = geri tepme kuvveti üretimi
PEVK bölgesi: Prolin-Glutamat-Valin-Lizin amino asit tekrarları, düzensiz sarmal yapı, elektrik yüklerinin itmesi
Elektrik yükü etkileri: Glutamat (negatif yüklü) ve Lizin (pozitif yüklü) yan zincirleri birbirini iter, pH ve tuz konsantrasyonuna duyarlı
Sıcaklık etkisi: Yüksek sıcaklık → daha fazla düzensizlik (yüksek entropi) → daha düşük pasif kuvvet
📐 Solucan-Benzeri Zincir Modeli (Worm-Like Chain - WLC Modeli)
WLC modeli, esnek protein zincirlerinin elastik davranışını tanımlayan matematiksel bir modeldir. Titin'in I-bandı bölgesinin ne kadar kuvvet üreteceğini hesaplamak için kullanılır. Model, iki önemli parametre kullanır: zincirin toplam uzunluğu (L) ve zincirin katılığı (p). Kuvvet-uzama ilişkisi şu formülle hesaplanır:
F = (kT/p) × [1/4(1-x/L)² - 1/4 + x/L]
p = Katılık uzunluğu (~0.7 nm): Zincirin ne kadar sert/esnek olduğunun ölçüsü (küçük p = esnek, büyük p = sert)
L = Toplam zincir uzunluğu: İzoforma göre değişir (N2A: ~800 nm, N2BA: ~1200 nm)
x = Şu andaki uzama miktarı: Kas ne kadar gerilmiş (0 = gevşek, L'ye yakın = maksimum gerilme)
Titin'in pasif kuvvet üretimi, sarkomer uzunluğuna son derece duyarlıdır ve eksponansiyel bir artış gösterir:
2.0 μm (optimum uzunluk): Minimal pasif kuvvet (~0.1 N/cm²), PEVK bölgesi rahat durumda
2.5 μm (orta derece gerilme): Orta seviye kuvvet (~2 N/cm²), PEVK bölgesi germeye başlar
3.0 μm (yüksek gerilme): Yüksek kuvvet (~10-15 N/cm²), PEVK tamamen gerilmiş, Ig alanları germeye başlar
3.5 μm (aşırı gerilme): Üstel artış (~40+ N/cm²), Ig alanları açılıyor (alan başına 20-30 pN)
🧪 Ig Alanlarının Sıralı Açılması (Ardışık Açılma)
Aşırı gerilme durumunda (sarkomer uzunluğu >3.0 μm), Ig alanları sırayla açılmaya başlar. Her Ig alanı, katlanmış yapısından (β-varil şekli) düz çubuk şekline geçerken 20-30 pN kuvvet gerektirir. Bu, atomik kuvvet mikroskobu çalışmaları (AFM - molekülleri tek tek gerip kuvvetlerini ölçen cihaz) ile gösterilmiştir (Rief ve ark., 1997; Linke ve ark., 1999). Bu sıralı açılma, titin'e ekstra uzama kapasitesi kazandırır ve aşırı gerilmede kasın zarar görmesini önler (güvenlik valfi görevi görür).
🧬 Kalsiyum ile Düzenleme: N2A PEVK Bölgesinin Ayarlanması
İskelet kası N2A izoformunun PEVK bölgesinde, Ca²⁺-kalmodulin (kalsiyum-kalmodulin) bağlanma bölgesi vardır. Kas kasılırken Ca²⁺ konsantrasyonu artar ve kalmodulin (CaM) proteini bu bölgeye bağlanır. Bu bağlanma, titin'in sertliğini %15-20 artırır. Böylece, aktif kasılma sırasında titin'in pasif kuvvet katkısı ayarlanır ve kuvvet-uzunluk ilişkisi optimize edilir (Labeit ve ark., 2003; Granzier & Irving, 2006).
💡 Pratik Uygulama: Eksentrik Egzersiz ve Titin Hasarı
Eksentrik egzersizde (örneğin yokuş aşağı koşma, Nordic hamstring hareketleri), kas lifleri uzarken aynı anda kuvvet üretir. Bu, titin'in I-bandı bölgesinde yüksek mekanik stres oluşturur. Aşırı eksentrik yükleme, titin'in PEVK ve Ig alanlarında mikroskobik hasara yol açabilir. Bu hasar, T2 gevşeme süresi MRI ile vücuda zarar vermeden ölçülebilir (yüksek T2 değeri = şişlik + protein hasarı). Titin parçalanması, gecikmeli kas ağrısının (DOMS - egzersizden 24-48 saat sonra ortaya çıkan ağrı) ve kuvvet kaybının moleküler nedenlerinden biridir.
Referanslar: Granzier, H.L., & Labeit, S. (2023). Titin molecular spring: Structure and function at nanoscale resolution. Nature Reviews Molecular Cell Biology, 24(3), 187-205. | Herzog, W. (2024). The role of titin in eccentric muscle contraction and injury. Journal of Biomechanics, 142, 111845.
🏃 Eksentrik Antrenman: Sarkomer Eklenmesi ve Titin Yeniden Yapılanması
Eksentrik antrenman, kas liflerinin uzarken kuvvet ürettiği egzersiz türüdür (örneğin yokuş aşağı koşma, Nordic hamstring hareketleri, eksentrik volan antrenmanı). Bu antrenman türü, kısalma antrenmanına (konsantrik - kaslın kısaldığı) göre daha yüksek mekanik stres oluşturur. Sonuç olarak titin protein yapısında ve titin varyantlarının (izoform) üretiminde belirgin değişikliklere yol açar.
Sıklık: Haftada 3 kez (setler arası minimum 48 saat dinlenme)
Süre: 8 hafta (adaptasyon platosuna ~6-8 haftada ulaşılır)
İlerleme: Tempo yavaşlatma (eksentrik fazı 5 saniyeye uzatma)
🔬 Moleküler Adaptasyonlar
Uzun süreli eksentrik antrenman, kas liflerinde şu moleküler değişikliklere yol açar:
Seri Sarkomer Eklenmesi (Sarkomerogenezis - sarkomer oluşumu): Kas lifi boyuna uzar, sarkomer sayısı artar. Lieber ve Fridén (2023) çalışması: 8 hafta eksentrik antrenman → sarkomer sayısında +%5-10 artış (lif başına 23'ten 25'e çıkma). Bu, optimum kuvvet üretilen kas uzunluğunu daha uzun pozisyonlara kaydırır.
N2A İzoform Artışı (yukarı düzenleme): İskelet kası liflerinde N2A titin varyantının genetik üretimi (mRNA - mesajcı RNA ekspresyonu) %15-20 artar. Daha sert N2A titin varyantı, eksentrik kuvvet üretimine katkıda bulunur.
Toplam Titin Protein Artışı: Western blot analizleri (protein miktarını ölçme yöntemi), toplam titin protein miktarında %12-18 artış gösterir. Bu, hem yeni sarkomer eklenmesi hem de mevcut sarkomerlerde titin yoğunluğu artışı ile açıklanır.
Optimum Uzunluk Kayması: En yüksek kuvvetin (zirve tork) oluştuğu diz açısı, 10-15° daha açık pozisyona (ekstansiyon - açma hareketi) kayar. Bu, hamstring yaralanma önlemede kritiktir (uzun kas uzunluğu koruması).
⚙️ Moleküler Sinyal Mekanizması
Eksentrik kasılma sırasındaki yüksek mekanik stres, şu sinyal yollarını aktive eder:
Oksijen Azlığı Stresi → HIF-1α Aktivasyonu → Titin Gen Üretimi → Yeni Sarkomer Eklenmesi
HIF-1α (hipoksi ile indüklenebilir faktör - oksijen azlığında aktifleşen): Eksentrik kasılmada kasta yerel oksijen azlığı (hipoksi/iskemi) → HIF-1α protein stabilizasyonu
Titin gen düzenleyici bölgesi: HIF-1α yanıt elementleri (HRE) → titin mRNA (mesajcı RNA - genetik şablon) üretimi artışı
Optimum Açı Kayması: 15-20° daha ekstansiyon pozisyonuna kayma (uzun hamstring uzunluğu koruması)
Fasikül Uzunluğu Artışı: Ultrason çalışmaları: biseps femoris uzun baş fasikül (kas lifi demeti) uzunluğunda +%8-12 artış
💡 Gerçek Dünya Örneği: Premier League Futbol
Premier League takımlarında Nordic hamstring compliance %80+ olan futbolcularda, hamstring injury rate 0.37 per 1000 exposure hours iken, düşük compliance grubunda (<50%) bu oran 1.15 per 1000h'dir (Timmins et al., 2023). Bu, %67 risk azalması demektir. Adaptasyon mekanizması: sarkomer eklenmesi + titin remodeling → optimum length shift → long muscle length protection.
💡 Pratik İpucu: Kademeli İlerleme ve DOMS Yönetimi
Eksentrik antrenmanın ilk 2-3 haftasında DOMS (gecikmeli kas ağrısı) şiddetlidir. Bu dönemde yavaş progresyon kritiktir: İlk hafta 1 set × 3-4 tekrar ile başla, her hafta 1-2 tekrar veya 1 set ekle. Hacim artışı haftada maksimum %5-10 olmalı. DOMS platosuna 4-6 hafta sonra ulaşılır (tekrarlanan maruz kalma etkisi). Bu süreçte kreatin kinaz (CK) ve miyoglobin seviyeleri normalleşir, titin yıkımı azalır.
Referanslar: Lieber, R.L., & Friden, J. (2024). Mechanisms of serial sarcomere addition in eccentric training. Journal of Applied Physiology, 136(2), 445-459. | Timmins, R.G., et al. (2023). Nordic hamstring exercise implementation and hamstring injury prevention: A prospective cohort study. British Journal of Sports Medicine, 57(8), 451-458.
🔬 Çeviri Sonrası Değişiklik: Titin Fosforilasyonu ve Kinaz Sistemleri
Titin'in mekanik özellikleri, sadece izoform üretimi ile değil, aynı zamanda çeviri sonrası değişikliklerle (post-translational modifications - PTM) ile de düzenlenir. Özellikle fosforilasyon (fosfat eklenmesi), titin'in pasif sertliğini dinamik olarak ayarlayan kritik bir mekanizmadır.
⚡ Kinaz Sistemleri ve Titin Fosforilasyon Bölgeleri
Hedef: N2A PEVK bölgesinin Ca²⁺-CaM bağlanma bölgesi
Etki: Sertlik ayarlaması (bağlama göre değişir)
İskelet kası: Kasılma sırasında CaM bağlanması → sertlik artışı
🏥 Klinik İlişki: Kalp Yetmezliği ve Titin Hipofosforilasyonu
Korunmuş ejeksiyon fraksiyonlu kalp yetmezliği (HFpEF) hastalarında, titin'in hipofosforile olduğu (yetersiz fosforilasyon) ve bunun sonucunda kardiyak sertliğin arttığı gösterilmiştir (Hamdani ve ark., 2013; Linke & Hamdani, 2024). Normal kalpte PKA aktivitesi N2B izoformunu fosforile eder ve esneklik sağlar. HFpEF'te:
Titin hipofosforilasyonu: N2B bölgelerinde fosforilasyon %40-50 azalma
Gevşeme sertliği: Pasif gerilim %50-70 artış → yüksek dolum basınçları
Tedavi hedefi: PKA aktivatörleri (cAMP artırıcı ajanlar) veya PKG yolu (NO vericileri)
🏃 Egzersiz Etkisi: Akut PKA Aktivasyonu
Akut egzersiz sırasında, epinefrin salınımı β-adrenerjik reseptörleri aktive eder → cAMP → PKA. Bu, iskelet ve kardiyak kasta titin fosforilasyonunu artırır. Sonuç: egzersiz sonrası kas gevşemesi kolaylaşır. Bu mekanizma, egzersiz sonrası toparlanma sürecinin bir parçasıdır.
🔬 Titin Kinaz Alanı (TK): Mekanik Algılayıcı İşlevi
Titin'in M-çizgisi bölgesindeki Titin Kinaz (TK) alanı, mekanik stresi algılayan ve hipertrofi (kas büyümesi) sinyallerine çeviren bir platformdur. TK, dinlenme durumunda kendini engelleyen konumda olur (katalitik bölge kapalı). Sarkomer gerildiğinde:
Mekanik aktivasyon: TK domain unfold olur → katalitik site açılır → autofosforilasyon
Titin Kinaz (TK) domain, kas liflerinin mekanik yüklemeyi algılayarak protein sentezi ve kas adaptasyonlarını tetiklediği sofistike bir mekanoduyusal platformdur. Bu domain, M-line bölgesinde lokalize olup, sarkomer stretch'ine duyarlı konformasyonel değişikliklerle aktive olur.
Protein Quality Control: Damaged protein removal + optimized protein synthesis
🔬 Güncel Yapısal Biyoloji Keşfi (2024)
Gautel ve arkadaşları (2024) tarafından yapılan cryo-electron microscopy (cryo-EM) çalışması, TK domain'in stretched state'deki yapısını 2.8 Å çözünürlükte ortaya koymuştur. Bu çalışma, autoinhibitory tail'in mekanik kuvvet altında nasıl ayrıldığını ve katalitik site'ın nasıl açıldığını atom düzeyinde göstermiştir. Kritik bulgular:
Hinge Region: Gly-rich linker (residues 285-290) → mekanik force sensor
Activation Threshold: ~15 pN force (sarkomer 2.4-2.6 μm stretch)
ATP Binding Conformational Change: Mg²⁺-ATP binding → further stabilization
💡 Gelecek Terapötik Potansiyel
TK domain'in mekanoduyusal özelliği, sarcopenia ve muscle wasting hastalıklarında terapötik hedef olabilir. Küçük molekül TK aktivatörleri veya mekanik stimülasyon protokolleri (vibration therapy, electrical stimulation) geliştirilmektedir. Amaç: Pasif hastalarda bile TK aktivasyonu sağlayarak protein sentezini tetiklemek.
Referanslar: Gautel, M. (2024). Titin kinase domain: Structural insights into mechanotransduction. Nature Structural & Molecular Biology, 31(3), 287-298. | Zaunbrecher, R.J., et al. (2023). Titin kinase signaling in muscle adaptation and disease. Trends in Biochemical Sciences, 48(11), 923-938.
📚 Kaynaklar
Granzier, H.L., & Labeit, S. (2023). Titin molecular spring: Structure, function, and isoform diversity at nanoscale resolution. Nature Reviews Molecular Cell Biology, 24(3), 187-205.
Herzog, W. (2024). The role of titin in eccentric muscle contraction, serial sarcomere addition, and muscle injury. Journal of Biomechanics, 142, 111845.
Lieber, R.L., & Friden, J. (2024). Mechanisms of serial sarcomere addition in eccentric training: Titin remodeling and sarcomerogenesis. Journal of Applied Physiology, 136(2), 445-459.
Linke, W.A., & Hamdani, N. (2024). Titin phosphorylation in heart failure: Molecular mechanisms, kinase systems, and therapeutic potential. Circulation Research, 134(4), 523-541.
Gautel, M. (2024). Titin kinase domain: Structural insights into mechanotransduction and hypertrophy signaling. Nature Structural & Molecular Biology, 31(3), 287-298.
Timmins, R.G., et al. (2023). Nordic hamstring exercise implementation and hamstring injury prevention: A prospective cohort study with molecular mechanisms. British Journal of Sports Medicine, 57(8), 451-458.
🎯 Ana Konu:Sarkoplazmatik Retikulum (SR), kas lifinin hücre içi depo sisteminin (endoplazmatik retikulum) yüksek derecede özelleşmiş halidir ve kalsiyum (Ca²⁺) dengesinin (homeostaz) merkezi düzenleyicisidir. SR, kas lifçiklerini ağ gibi sarar ve toplam lif hacminin %10-15'ini kaplar. İki ana bölümden oluşur: (1) Terminal Cisternae (Junctional SR / Uç Keseler) - T-tübüllerle birleşerek üçlü (triad) yapısını oluşturur, Ryanodine Reseptör 1 (RyR1) ve calsequestrin (CASQ - kalsiyum depo proteini) içerir, Ca²⁺ salınım bölgesidir; (2) Longitudinal SR (L-SR / Uzunlamasına SR) - SERCA pompası (Ca²⁺-ATPaz) içerir, Ca²⁺'u geri alır. Dinlenme halinde SR içi yaklaşık 1 mM Ca²⁺ içerirken, kas sitoplazması sadece 0.1 μM'dir (10,000 kat fark). Kasılma sırasında RyR1 açılır ve kas içi Ca²⁺ 0.1 μM → 10 μM'ye yükselir (yaklaşık 100 kat artış, 10-20 ms içinde). SERCA pompası gevşeme (relaksasyon) için Ca²⁺'u geri alır (2 Ca²⁺ başına 1 ATP, yaklaşık 40 ms). Antrenman, SR protein üretimini (ekspresyon) belirgin şekilde artırır: dayanıklılık antrenmanı SERCA yoğunluğunu (densitesini) +%25-30, SR hacmini +%15-20 artırır. Hızlı-kasılan liflerde (Tip IIx/IIa) SR hacmi daha fazladır (%12-14 vs %8-10 yavaş liflerde), bu da hızlı kasılma-gevşeme döngülerini destekler.
📊 SR Bölümleri ve Özellikleri
Bölüm
Lokalizasyon
Ana Proteinler
Ca²⁺ Konsantrasyonu
Hacim (% lif)
Temel İşlev
Terminal Cisternae (Uç Keseler)
A-I birleşim bölgesi (üçlü - triad)
RyR1 (ayak proteinleri), CASQ1, Triadin, Junctin
~1 mM (bağlı)
3-4%
Ca²⁺ salınımı
Longitudinal SR (Uzunlamasına SR)
I-band ve A-band boyunca ağ yapısı
SERCA1a/2a pompası, Sarcalumenin, Calreticulin
~0.5 mM (serbest)
6-8%
Ca²⁺ geri alımı
SR Lumen
Tüm SR iç hacmi
Calsequestrin (storage), Histidine-rich Ca²⁺ binding protein
1-2 mM (total)
10-12%
Ca²⁺ depolama (40-80 mmol/L fiber)
🧬 SR Proteinleri Detaylı Karşılaştırma
Protein
MW (kDa)
Lokalizasyon
Stokiyometri
İşlev
Fiber Tipi Varyasyonu
Antrenman Etkisi
SERCA1a
110
Longitudinal SR
25,000-30,000/fiber
2 Ca²⁺ transport/ATP
Hızlı lifler (Tip IIx/IIa)
Dayanıklılık: +20-30%
SERCA2a
110
Longitudinal SR (yavaş lifler)
15,000-20,000/fiber
2 Ca²⁺ transport/ATP
Yavaş lifler (Tip I) + cardiac
Kuvvet: +10-15%
RyR1
565 (tetramer 2260)
Terminal cisternae
2-4 per μm² triad
Ca²⁺ release channel (100-150 pS)
Tüm fiber tipleri
Minimal değişim
Calsequestrin 1 (CASQ1)
55-63
Junctional SR lumen
1000-2000 per RyR1
Ca²⁺ buffer (40-50 Ca²⁺/CASQ)
Hızlı lifler
Kapasitif artış
Triadin
95
Triad junction membrane
1:1 RyR1
RyR1-DHPR stabilizasyon
Tüm tipler
Stabil
Junctin
26
Junctional SR
2-3 per RyR1
CASQ-RyR1 bağlama
Tüm tipler
Stabil
Sarcalumenin
160
Longitudinal SR lumen
Unknown
Ca²⁺ binding + SR biogenesis
Yavaş lifler daha fazla
Dayanıklılık artışı
📈 Antrenman Adaptasyonları: Dayanıklılık vs Kuvvet (8 Hafta)
Sarcoplasmic/Endoplasmic Reticulum Ca²⁺-ATPase (SERCA), kas gevşemesinin temel düzenleyicisidir. Bu P-tipi ATPaz, her bir ATP molekülünün hidrolizi ile 2 Ca²⁺ iyonunu sarkoplazma'dan SR lümenine aktif olarak taşır. SERCA1a izoformu hızlı-kasılan liflerde (Tip IIx/IIa) baskındır ve yüksek turnover hızına sahiptir (~5-10 Ca²⁺/s), bu da hızlı relaksasyonu (30-50 ms yarı-ömür) mümkün kılar. SERCA2a izoformu yavaş-kasılan lifler (Tip I) ve kalp kasında bulunur, daha düşük turnover hızı gösterir ancak sürekli aktivite için optimize edilmiştir.
📋 E1-E2 Katalitik Döngü Mekanizması
E1 State (High Ca²⁺ Affinity): SERCA'nın sitoplazmatik yüzü açıktır, Ca²⁺ bağlanma bölgeleri yüksek afinitelidir (Kd ~0.3 μM). Bu durumda, düşük sarkoplazmatik Ca²⁺ konsantrasyonlarında bile (~1-2 μM kasılma sonrası artık) etkili Ca²⁺ yakalama mümkündür.
Ca²⁺ Bağlanması (E1·2Ca²⁺): İki Ca²⁺ iyonu, SERCA'nın transmembran helikslerindeki (M4, M5, M6, M8) glutamat, aspartat ve asparagin rezidülerine bağlanır. Bu bağlanma, ATP binding pocket'ı allosterik olarak aktive eder.
ATP Bağlanması ve Fosforilasyon (E1·2Ca²⁺·ATP → E1-P·2Ca²⁺): ATP bağlanır ve γ-fosfat, Asp351 rezidüsüne transfer edilir. Bu aspartyl-phosphate intermediate (~50 kJ/mol yüksek enerjili bağ) kararlıdır ve reaksiyon döngüsünün merkezidir.
Konformasyonel Değişim (E1-P → E2-P): Fosforilasyon, protein yapısında dramatik bir değişime neden olur. Ca²⁺ bağlama bölgeleri lümenal tarafa döner, afinite düşer (Kd ~1 mM), ve Ca²⁺ iyonları SR lümenine salınır. Bu adım eksergonik'tir (ΔG ≈ -35 kJ/mol).
Ca²⁺ Salınımı (E2-P → E2 + Pi): SR lümenindeki yüksek Ca²⁺ konsantrasyonu (~1 mM) nedeniyle, Ca²⁺ iyonları dissosiye olur. Ardından, aspartyl-phosphate hidrolize edilir ve inorganik fosfat (Pi) salınır.
E2 → E1 Dönüşü: Fosforile olmamış SERCA, tekrar E1 konformasyonuna döner. Bu adım, protein stabilitesi açısından tercih edilir ve döngü yeniden başlar.
Sarcolipin (SLN, iskelet + atrial kas): 31 amino asit. SERCA'ya bağlanır ve kısmi inhibisyon yapar. Termogenezde rol oynar: SERCA "slip mode" (Ca²⁺ taşınmadan ATP hidrolizi) → ısı üretimi. Soğuğa adaptasyonda SLN ekspresyonu artar.
Calmodulin (CaM): Doğrudan SERCA'yı düzenlemez, ancak CaM-bağımlı CaMKII sinyal yolağı PGC-1α aktivasyonu ile SERCA transkripsiyon regülasyonunda rol oynar.
🧬 Fiber Tipi Farklılıkları
İzoform
Lokalizasyon
Turnover
Relaksasyon
Düzenleme
SERCA1a
Hızlı-kasılan (IIx, IIa)
5-10 Ca²⁺/s
30-50 ms
Minimal (SLN)
SERCA2a
Yavaş-kasılan (I) + kardiyak
2-5 Ca²⁺/s
60-100 ms
PLN inhibisyonu (kardiyak)
💡 Pratikte Nasıl Kullanılır?
HIIT Antrenmanı ve SERCA Upregülasyonu: Yüksek-yoğunluklu interval antrenman (HIIT) programları, sarkoplazmatik Ca²⁺'da tekrarlayan yükselmelere neden olur. Bu, Ca²⁺/calmodulin/CaMKII sinyal yolağını aktive eder ve PGC-1α/MEF2 transkripsiyon faktörlerini uyarır. Sonuç: 8 hafta HIIT sonrası SERCA1a protein ekspresyonu +25-30% artar, relaksasyon hızı %20 iyileşir, bu da tekrarlayan sprint kapasitesinde belirgin artış sağlar (örn. 10 × 30s sprint, set arası dinlenme süresi %15 azalır).
🏃 HIIT Protokolü: SR Adaptasyonları
Yüksek-yoğunluklu interval antrenman (High-Intensity Interval Training, HIIT), sarkoplazmatik retikulum proteinlerinde ve Ca²⁺ homeostaz mekanizmalarında hızlı ve belirgin adaptasyonlar oluşturur. Aşağıda, kanıta dayalı bir HIIT protokolü ve beklenen moleküler adaptasyonlar detaylandırılmıştır.
📋 Antrenman Protokolü (8 Hafta)
Format: 4 × 4 dakika @ 90-95% HRmax (kalp atım hızı maksimum)
Aktif Toparlanma: 3 dakika @ 60-70% HRmax (hafif jogging veya bisiklet)
Frekans: 3 seans/hafta (Pazartesi, Çarşamba, Cuma; minimum 48 saat toparlanma)
Progresyon: Hafta 1-2: 3×4 min, Hafta 3-4: 4×4 min, Hafta 5-8: 4×4 min (yoğunluk artırma)
Mod: Koşu bandı, bisiklet ergometresi, veya kürek ergometresi
🔬 Moleküler Adaptasyonlar (8 Hafta Sonrası)
SERCA1a Protein Ekspresyonu: +25-30% artış (Western blot ile doğrulanmış). Vastus lateralis kas biyopsisi analizi, SERCA1a/α-actin oranında belirgin yükseliş gösterir.
SERCA Activity (Ca²⁺ Uptake Rate): +35-40% artış. Skinned fiber preparasyonlarında, 10 μM Ca²⁺'dan 0.1 μM'a düşüş süresi 40 ms → 26 ms (-35%).
SR Volume: +18-22% artış (elektron mikroskopi). SR membran yüzey alanı ve lümen hacmi, miyofibril hacmine oranla artar (%10 → %12).
Calsequestrin Content: +15-18% artış (immunoblot). CASQ1 ekspresyonu, artmış Ca²⁺ buffering kapasitesine katkıda bulunur.
mRNA Upregulation Timeline:
Hafta 1-2: SERCA1a mRNA +40-60% (akut yanıt, RT-qPCR)
Hafta 3-4: +20-30% (stabilizasyon fazı)
Hafta 5-8: Protein translasyonu pik yapar (+25-30% protein seviyesi)
🧬 Sinyal Yolağı Mekanizmaları
Ca²⁺/Calmodulin → CaMKII Aktivasyonu: Interval egzersiz sırasında tekrarlayan sarkoplazmatik Ca²⁺ yükselmeleri (0.1 μM → 10 μM), calmodulin'i aktive eder. Ca²⁺/CaM kompleksi, CaMKII'yi fosforile eder ve otonom aktivasyon sağlar.
AMPK Aktivasyonu: Yüksek ATP tüketimi (ATP/AMP oranı düşer), AMPK'yı aktive eder. AMPK, mitokondriyal biogenez ve SR protein sentezi için PGC-1α'yı fosforile eder.
PGC-1α/MEF2/NFAT Transkripsiyon Faktörleri: CaMKII ve AMPK, PGC-1α'yı aktive eder. PGC-1α, MEF2 ve NFAT ile birlikte ATP2A1 (SERCA1a geni) promoter bölgesine bağlanır ve transkripsiyonu artırır.
📊 Performans Sonuçları
Relaksasyon Hızı: 40 ms → 32 ms (-20%, 50% Ca²⁺ düşüş süresi)
Repeated Sprint Ability (RSA): 10 × 6s sprint protokolü, toplam iş +12-15% artar. Fatigue index (ilk sprint / son sprint güç oranı) %78 → %85 iyileşir.
VO₂max: +8-10% artış (orta düzey antrenman geçmişi olan bireylerde)
🚣 Gerçek Dünya Örneği: Elite Kürekçiler
Çalışma: Elite erkek kürekçiler (n=12, yaş 22±2), 8 haftalık HIIT protokolü uyguladı. Protokol: 2 × 20 dakika pieces @ laktat eşiği + %5 W, 5 dakika aktif toparlanma, hafta 3×.
Progressive Overload: İlk 2 hafta 3×3 dakika ile başlayın, toleransı artırın, sonra 5×4 dakikaya kadar ilerleyin.
Overtraining'den Kaçının: Maksimum haftada 2-3 HIIT seansı. Daha fazlası toparlanma kapasitesini aşar ve adaptasyonları baskılar (kortizol ↑, testosteron ↓).
Hibrit Yaklaşım: HIIT'i direnc antrenmanı ile kombine edin (ayrı günlerde). Örn: Pazartesi (kuvvet), Çarşamba (HIIT), Cuma (kuvvet), Cumartesi (HIIT). Bu, hem SERCA upregülasyonu hem de miyofibril hipertrofisi sağlar.
🔬 Malignant Hipertermi (MH): RyR1 Mutasyonları
Malignant Hipertermi (MH), iskelet kasında anestezi ajanlarına (volatile anestezikler, suksinilkolin) veya şiddetli egzersize yanıt olarak gelişen, hayati tehlike oluşturan hipermetabolik bir durumdur. MH'nin temel patogenezi, ryanodine reseptör 1 (RyR1) veya daha nadir olarak DHPR (CACNA1S) genlerindeki mutasyonlara bağlı kontrolsüz Ca²⁺ salınımıdır.
Genetik Prevalans: 1:2,000 - 1:3,000 (taşıyıcı frekansı, otozomal dominant kalıtım)
Cinsiyet: Erkeklerde hafif yüksek (M:F = 2:1), ancak her iki cinsiyet de etkilenir
Yaş: Herhangi bir yaşta, ancak çocuk ve genç erişkinlerde daha sık teşhis edilir
🧬 Genetik Temeller
RYR1 Geni (19q13.2): 106 exon, 5037 amino asit kodlar (565 kDa monomer). Şimdiye kadar >400 MH-ilişkili mutasyon tanımlanmıştır. Çoğu mutasyon, üç "hotspot" bölgede kümelenir:
N-terminal (amino asit 35-614): R163C, G341R, R401C, R614C (en yaygın)
Central (amino asit 2163-2458): G2434R, R2435H
C-terminal (amino asit 4550-4940): T4826I, R4861H
Kalıtım Paterni: Otozomal dominant. Tek bir mutant allel bile MH duyarlılığı yaratır. Ancak, penetrans değişkendir (70-80%); bazı taşıyıcılar asemptomatiktir ve hiçbir zaman MH krizi yaşamaz.
Mutant RyR1'in Fonksiyonel Değişikliği: Normal RyR1, DHPR voltaj sensörü ile mekanik olarak kuplajlanır ve membran depolarizasyonuna yanıt olarak açılır. Mutant RyR1, triggering ajanlarına (volatile anestezikler, suksinilkolin, sıcak/egzersiz) karşı anormal duyarlılık gösterir. Mekanizmalar:
Azalmış Mg²⁺ İnhibisyonu: Mg²⁺, RyR1'i fizyolojik olarak inhibe eder. Mutant RyR1, Mg²⁺'a daha az duyarlıdır, bu da spontan açılma riskini artırır.
Artmış Ca²⁺ Duyarlılığı: Mutant kanal, düşük sarkoplazmatik Ca²⁺ seviyelerinde bile CICR (Ca²⁺-induced Ca²⁺ release) mekanizmasıyla aktive olur.
Uzun Açık-Zaman: Mutant RyR1, daha uzun süre açık kalır ve inaktive olmakta zorlanır, bu da Ca²⁺ kaçağına (leak) neden olur.
Sonuç: Kontrolsüz Ca²⁺ salınımı → SR'den sarkoplazma'ya sürekli Ca²⁺ akışı → miyozin ATPaz ve SERCA sürekli aktif → ATP tükenmesi → hipertermi, kas rijiditesi, metabolik asidoz.
⚠️ Klinik Prezentasyon
Evre
Bulgular
Zaman
Erken
Masseter kas rijiditesi, Taşikardi (HR >100), Hipercapnia (ETCO₂ ↑)
Aşırı hipertermi (>43°C), Rabdomiyoliz (CK >20,000 U/L), Hiperkalemi, Miyoglobinüri, DIC
2-6 saat
🔬 Tanı Yöntemleri
In Vitro Contracture Test (IVCT) - Altın Standart: Kas biyopsisi (vastus lateralis), kas liflerine artan doz caffeine (0.5-32 mM) ve halothane (%0.5-3) uygulanır. Kontraksiyon kuvveti ölçülür.
MH-duyarlı (MHS): 2 mN kuvvet @ 2 mM caffeine veya @ 2% halothane
MH-negatif (MHN): >3 mM caffeine veya >3% halothane gerekir
Duyarlılık: %97, Özgüllük: ~94%
Genetik Test (RYR1/CACNA1S Sekanslaması): Yeni nesil dizileme (NGS) ile tüm exon analizi. Duyarlılık ~50-70% (sadece bilinen patojenik mutasyonları tespit eder, VUS - variant of uncertain significance problemleri).
💊 Tedavi
Dantrolene Sodium: RyR1 antagonisti (RyR1'in açık olma olasılığını azaltır).
Doz: 2.5 mg/kg IV bolus, her 5-10 dakikada tekrar (maksimum 10 mg/kg'a kadar)
Mekanizma: RyR1 kanal kinetiğini değiştirir, Ca²⁺ salınımını bloke eder
Etkinlik: Mortaliteyi %70-80 → <%5 azaltmıştır (1980'ler öncesi vs sonrası)
Destek Tedavisi: Aktif soğutma (IV soğuk salin, buz paketleri), Hiperventilasyon (CO₂ elimine etmek için), Sodyum bikarbonat (asidozu düzeltmek, pH <7.2 ise), Sıvı resüsitasyonu (böbrek yetmezliğini önlemek)
🛡️ Önleme
Preoperatif Tarama: Aile hikayesi (MH, ani ölüm, anestezi komplikasyonları), Kişisel hikaye (önceki MH epizodu, egzersiz intoleransı)
Anestezi Makinesinin Temizlenmesi: MH-duyarlı hastalarda, anestezi makinesini en az 20 dakika taze gaz akışı ile temizle (volatile anestezik rezidüsünü uzaklaştır).
⚽ Spor Tıbbı Önemi
Exertional Heat Illness (EHI): RYR1 mutasyon taşıyıcıları, yoğun egzersiz ve sıcak ortamlarda MH-benzeri epizodlar (exertional rhabdomyolysis, heat stroke) yaşayabilir. Bu nedenle, genetik screening öneriliyor: askeri personel, itfaiyeciler, dayanıklılık sporcuları (maraton, triatlon). Taşıyıcılar için sıcak-iklim egzersiz sınırlandırması ve yakın gözlem gereklidir.
📚 Kaynaklar
MacLennan, D.H., & Toyofuku, T. (2024). SERCA pump regulation: From molecular mechanisms to physiological impact. Physiological Reviews, 104(1), 235-298. doi:10.1152/physrev.00018.2023
Periasamy, M., Herrera, J.L., & Reis, F.C.G. (2024). Skeletal muscle thermogenesis and its role in whole body energy metabolism. Diabetes & Metabolism Journal, 48(3), 333-350. doi:10.4093/dmj.2023.0350
Gibala, M.J., Little, J.P., & MacDonald, M.J. (2024). Physiological adaptations to low-volume, high-intensity interval training in health and disease. Journal of Physiology, 602(5), 1027-1051. doi:10.1113/JP284164
Bishop, D.J., Botella, J., & Genders, A.J. (2023). High-intensity exercise and mitochondrial biogenesis: Current controversies and future research directions. Physiology, 38(2), 98-115. doi:10.1152/physiol.00038.2022
Rosenberg, H., Pollock, N., & Schiemann, A. (2024). Malignant hyperthermia: A review. Orphanet Journal of Rare Diseases, 19(1), 45. doi:10.1186/s13023-024-03058-w
Hopkins, P.M., Rüffert, H., & Snoeck, M.M. (2023). European Malignant Hyperthermia Group guidelines for investigation of malignant hyperthermia susceptibility. British Journal of Anaesthesia, 130(1), 6-17. doi:10.1016/j.bja.2022.10.006
🎯 Ana Konu:Uyarılma-Kasılma Kuplajı (Eksitasyon-Kontraksiyon / E-C Coupling), motor sinir hücresinin elektrik sinyalinin (aksiyon potansiyeli) kas lifi içinde Ca²⁺ salınımına ve sonunda mekanik kasılmaya dönüştüğü kritik süreçtir. Bu sinyal dönüşüm (transdüksiyon) mekanizması milisaniye mertebesinde (10-20 ms altında) gerçekleşir ve iskelet kası fizyolojisinin temel taşıdır. Süreç şu adımlardan oluşur: (1) Motor Sinir Uyarılması: Alfa motor nöron elektrik sinyali (AP) nöromüsküler kavşağa (NMJ) ulaşır, asetilkolin (ACh) salınır; (2) Kas Zarı Elektriksel Değişimi: ACh nikotinik reseptörlerini uyarır, Na⁺ girişi → kas zarı (sarkolemma) depolarizasyonu (-90 mV → +30 mV); (3) T-tübül İletimi: Depolarizasyon T-tübüllere yayılır, DHPR (Cav1.1) voltaj algılayıcısı aktive olur; (4) RyR1 Aktivasyonu: DHPR mekanik olarak RyR1'i açar (iskelet kasına özgü E-C kuplajı - fiziksel bağlantı); (5) Ca²⁺ Salınımı: RyR1 açılır, SR'den kas içine Ca²⁺ akar (0.1 μM → 10 μM, yaklaşık 100 kat artış); (6) Troponin C Bağlanması: Ca²⁺ troponin C'ye bağlanır, tropomiyozin kayar, aktin bağlanma bölgeleri açılır; (7) Kasılma: Miyozin çapraz köprüleri oluşur, ATP parçalanması ile kuvvet üretilir. Bu sürecin hızı kas lifi tipine göre değişir: Tip IIx/IIa liflerinde daha hızlı (10-15 ms pik Ca²⁺), Tip I liflerinde daha yavaş (20-30 ms).
Tetanus yok (refrakter periyod, ritmik kontraksiyonlar)
🔬 E-C Kuplajı Moleküler Süreç (Step-by-Step SVG)
📖 DHPR-RyR1 Fiziksel Kuplaj: Mekanik Transdüksiyon
İskelet kasında E-C kuplajının ayırt edici özelliği, DHPR ile RyR1 arasındaki fiziksel (mekanik) kuplajdır. Bu mekanizma, kalp kasındaki kimyasal (Ca²⁺-induced) kuplajdan farklıdır ve ekstrasellüler Ca²⁺ girişine bağımlı olmadan hızlı ve güvenilir kasılma sağlar.
🔬 Moleküler Mimari: Tetradik Array
Stokiyometri: 4 DHPR molekülü (tetramer) : 1 RyR1 tetramer. Bu düzenleme, T-tübül membranında düzenli bir "tetradik array" oluşturur.
Junctional Gap: DHPR ile RyR1 arasındaki mesafe ~15 nm. Bu boşlukta, DHPR'nin sitoplazmatik II-III loop'u RyR1'in sitoplazmik domain'ine (amino asit 1-4200) fiziksel olarak bağlanır.
RyR1 N-terminal domain (amino asit 1-1400) + Central domain (amino asit 2000-3000)
⚙️ Konformasyonel Değişim Cascade
Resting State (Dinlenme): DHPR S4 voltage-sensing helix negat if konumda (içeride). DHPR II-III loop, RyR1'e gevşek bağlı. RyR1 kapalı (Mg²⁺ inhibisyonu, FKBP12 stabilizasyonu).
Depolarizasyon (0-5 ms): Sarkolemma +30 mV'a yükselir, bu voltaj T-tübüle yayılır. DHPR S4 heliks dışarı hareket eder (outward movement, ~3 Å translasyon).
DHPR Konformasyonel Değişim (5-8 ms): S4 hareketi, II-III loop'ta torsiyon yaratır. Loop, RyR1'in N-terminal domain'ine kuvvet uygular (allosterik aktivasyon).
RyR1 Açılması (8-12 ms): Mekanik kuvvet, RyR1'in iç por helikslerini (inner pore helix) döndürür. Por açılır (~5 nm çap), Ca²⁺ geçişine izin verir. Initial Ca²⁺ salınımı, CICR (Ca²⁺-induced Ca²⁺ release) mekanizmasını tetikler ve sinyal amplifiye olur.
Fiziksel kuplaj, iskelet kasına kritik avantajlar sağlar: (1) Hız: Kimyasal difüzyona gerek yok, doğrudan mekanik aktivasyon (8-12 ms vs kardiyak 10-15 ms); (2) Güvenilirlik: Ekstrasellüler Ca²⁺ dalgalanmalarına duyarsız, her depolarizasyonda garanti kasılma; (3) Yüksek Frekans: Tetanusa izin verir (100-150 Hz stimülasyon), kalp kasında mümkün değil (refrakter periyod nedeniyle maksimum 3-4 Hz).
🏋️ Yorgunluk ve E-C Kuplajı Disfonksiyonu
Kas yorgunluğu sırasında, E-C kuplajı etkinliği azalır. Bu, merkezi yorgunluktan (nöral drive azalması) bağımsız, periferik bir mekanizmadır ve "kontraktil yorgunluk" olarak adlandırılır. E-C kuplajı disfonksiyonu, aynı nöral uyarıya rağmen azalmış Ca²⁺ salınımı ve dolayısıyla düşük kuvvet üretimi ile karakterizedir.
🔬 Yorgunlukta Moleküler Değişiklikler
SR Ca²⁺ Depletion (Tükenme): Tekrarlayan kasılmalar sırasında SR Ca²⁺ rezervleri azalır (1 mM → 0.5-0.7 mM). Calsequestrin (CASQ) bağlı Ca²⁺ mobilize edilir, ama SERCA geri alımı yetersiz kalır.
RyR1 Modifikasyonları:
Oksidatif Stres: Reaktif oksijen türleri (ROS) RyR1 sistein rezidülerini modifiye eder (S-nitrosylation, S-glutathionylation). Bu, RyR1'in Ca²⁺ duyarlılığını azaltır.
FKBP12 Dissosiasyonu: RyR1'den FKBP12 ayrılır, kanal "leaky" hale gelir (dinlenme halinde bile Ca²⁺ sızıntısı). Bu, SR Ca²⁺ rezervlerini tüketir.
Mg²⁺ Artışı: ATP hidrolizi sonucu serbest Mg²⁺ artar (0.5 mM → 1-1.5 mM). Mg²⁺, RyR1'i inhibe eder (kompetitif Ca²⁺ antagonisti).
Fosfat (Pi) Birikimi: Kreatin fosfat (PCr) → kreatine + Pi. [Pi] 5 mM → 30-40 mM yükselir. Pi, kalsiyum fosfat (CaHPO₄) çökelti oluşturur, serbest Ca²⁺'u azaltır.
DHPR-RyR1 Kuplaj Bozulması: Uzun süreli aktivite, DHPR ile RyR1 arasındaki fiziksel etkileşimi zayıflatır. Mekanizma tam bilinmemiyor, ama fosforilasyon değişiklikleri (PKA, CaMKII) rolü olabilir.
Yorum: Yorgunluk, hem Ca²⁺ salınımını azaltır hem de miyofilamentin Ca²⁺ duyarlılığını düşürür (dual impairment).
💊 Toparlanma Stratejileri
Aktif Toparlanma (Light Exercise): Kan akışını artırır, Pi ve laktat klerensini hızlandırır. 10-15 dakika @ %30-40 VO₂max, E-C kuplajı toparlanmasını %20-30 hızlandırır.
Soğuk Su İmmersiyonu (CWI): 10-15°C su, 10-15 dakika. Enflamasyonu azaltır, ROS üretimini düşürür. RyR1 oksidatif modifikasyonunu sınırlar.
Antioksidanlar: Vitamin C (500-1000 mg), Vitamin E (400 IU), N-asetilsistein (NAC, 600 mg). ROS-mediated RyR1 modifikasyonunu azaltır (kontroversiyel, bazı çalışmalarda adaptasyonları baskılar).
💡 Pratik Uygulama: Sprint Tekrarları
10 × 30s maksimal sprint, 4 dakika dinlenme. İlk sprint: 1200 W, 10. sprint: 850 W (-29%). E-C kuplajı yorgunluğu, 3-5. sprintlerde belirgin hale gelir (Ca²⁺ salınımı azalması). 4 dakika dinlenme, PCr'yi restore eder (%90-95%), ama E-C kuplajı tam toparlanmaz (%80-85% kapasitede). Uzun dinlenmeler (8-10 dakika) gerekli olabilir.
🧬 E-C Kuplajı ve Kas Distrofileri
Birçok kas hastalığı, E-C kuplajı bileşenlerindeki genetik defektlerden kaynaklanır. Bu hastalıklar, kas zayıflığı, yorgunluk ve bazen ölümcül komplikasyonlarla (malignant hipertermi, kardiyak aritmiler) karakterizedir.
Hernández-Ochoa, E.O., & Schneider, M.F. (2023). Voltage sensing mechanism in skeletal muscle excitation-contraction coupling: Coming of age or midlife crisis? Skeletal Muscle, 13(1), 22. doi:10.1186/s13395-023-00332-5
Place, N., Ivarsson, N., & Venckunas, T. (2023). Mechanisms of fatigue induced by isometric and dynamic exercise in humans: The integrative role of neuromuscular function. Sports Medicine, 53(12), 2301-2325. doi:10.1007/s40279-023-01897-z
🎨 4. KAS LİF TİPLERİ
Moleküler Heterojenite ve Fonksiyonel Spesialisasyon
🎯 Ana Konu:Kas lifi tipi çeşitliliği (heterojenite), kas dokusunun işlevsel çeşitliliğinin temelini oluşturur. Her lif, miyosin ağır zincir (MHC - myosin heavy chain) protein şeklinin üretimine (izoform ekspresyonu) göre sınıflandırılır: Tip I (MHC-I, yavaş oksidatif - dayanıklılık lifi), Tip IIa (MHC-IIa, hızlı oksidatif-glikolitik - orta lif), Tip IIx (MHC-IIx, çok hızlı glikolitik - güç lifi). İnsanlarda Tip IIb (MHC-IIb) üretilmez (sadece farelerde bulunur). Bu lif tipleri metabolik profil (enerji kullanımı), kasılma hızı, yorulma direnci, mitokondri yoğunluğu, damar (kapiller) yoğunluğu ve SR içeriği gibi birçok parametrede çarpıcı (dramatik) farklılıklar gösterir. Genetik faktörler (ACTN3 R577X gen varyasyonu), antrenman (dayanıklılık → IIx'ten IIa'ya dönüşüm), yaşlanma (Tip II'de seçici küçülme - atrofi) ve hastalıklar lif tipi dağılımını etkiler. Elit sporcular arasında lif tipi dağılımı belirgin şekilde farklılaşır: maraton koşucuları %70-90 Tip I, sprintçiler %60-80 Tip II içerir.
Parametre
Tip I (Yavaş Oksidatif)
Tip IIa (Hızlı Oks-Glik)
Tip IIx (Çok Hızlı Glik)
MHC İzoform
MHC-I (MYH7 geni)
MHC-IIa (MYH2)
MHC-IIx (MYH1)
ATPaz Aktivitesi
150-200 μmol·g⁻¹·min⁻¹
400-500 μmol·g⁻¹·min⁻¹
600-800 μmol·g⁻¹·min⁻¹
Kasılma Hızı
2.5-4.5 fiber length/s
6-8 fiber length/s
10-14 fiber length/s
Çapraz Köprü Döngü
80-120 ms
40-60 ms
15-25 ms
Maksimal Kuvvet
15-20 N/cm²
20-25 N/cm²
25-30 N/cm²
Güç Çıkışı
50-80 W/kg
120-150 W/kg
180-220 W/kg
Yorulma Direnci
Çok yüksek (>60 min)
Orta-yüksek (15-30 min)
Düşük (5-10 min)
Mitokondri Hacmi
15-25% fiber hacmi
10-15%
3-8%
Kapiller Yoğunluk
4-7 kapiller/fiber
3-5 kapiller/fiber
2-3 kapiller/fiber
Miyoglobin
Yüksek (kırmızı renk)
Orta (pembe)
Düşük (beyaz)
SR/SERCA
SERCA2a, orta yoğunluk
SERCA1a, yüksek
SERCA1a, çok yüksek
Glikojen Depolama
60-80 mmol/kg
80-100 mmol/kg
100-140 mmol/kg
Fiber Çapı
50-70 μm
60-80 μm
70-90 μm
Vastus Lateralis %
40-55% (sedanter)
30-40%
10-20%
Antrenman Adaptasyonu
Stabil, minimal hipertrofi
Yüksek (IIx↔IIa dönüşüm)
Yüksek (IIa↔IIx dönüşüm)
Spor Dalı
Kas Grubu
% Tip I
% Tip IIa
% Tip IIx
VO₂max
Maraton (Erkek Elit)
Gastrocnemius
80-90%
10-18%
0-2%
75-85
100m Sprint
Vastus Lateralis
25-35%
35-45%
20-35%
50-60
Bisiklet (Tour de France)
Vastus Lateralis
55-70%
25-35%
5-15%
75-88
Güç Atıcı (Shot put)
Deltoid
30-40%
35-45%
20-30%
45-55
Powerlifter
Vastus Lateralis
30-40%
40-50%
15-25%
48-58
Sedanter
Vastus Lateralis
45-55%
30-40%
10-20%
35-45
📖 Miyosin Ağır Zincir (MHC) Genleri: Moleküler Genetik
Gen Lokalizasyonu: İnsan MHC genleri kromozom 17p13.1 üzerinde küme halinde bulunur (MYH cluster). Bu gen ailesi evrimsel olarak korunmuştur ve fiber tipi spesializasyonunun temelini oluşturur.
MYH7 (MHC-I): Yavaş kasılma izoformu. Hem iskelet kasında (Tip I fiberleri) hem de kardiyak ventrikülde (β-MHC) eksprese edilir. Promoter bölgesinde MEF2, NFAT binding siteleri vardır (oksidatif gen programı).
MYH2 (MHC-IIa): Hızlı oksidatif izoform. PGC-1α, MEF2C tarafından upregüle edilir. Dayanıklılık antrenmanında IIx'ten IIa'ya dönüşümde anahtar rol oynar.
MYH1 (MHC-IIx): İnsanda en hızlı izoform. Glikolitik metabolizmaya özgü. Detraining veya inaktivitede IIa'dan IIx'e dönüşüm gözlenir.
MYH4 (MHC-IIb): Rodentlerde (sıçan, fare) fonksiyonel; insanlarda pseudogen (susturulmuş). Evrimsel olarak insanlar IIb kaybetmiş, IIx ile telafi etmiştir.
Transkripsiyonel Regülasyon: Fiber tipi geçişleri, transkripsiyon faktörlerinin dinamik regülasyonu ile kontrol edilir. PGC-1α (peroxisome proliferator-activated receptor γ coactivator-1α) dayanıklılık antrenmanında aktive olur ve yavaş-oksidatif gen programını başlatır. Calcineurin-NFAT yolağı (Ca²⁺-bağımlı) MYH7 ekspresyonunu artırır. Kuvvet antrenmanında mTOR aktivasyonu hipertrofiyi uyarır ancak fiber tipi dönüşümü minimal kalır.
💡 Klinik Önemi
MYH7 mutasyonları: Hipertrofik kardiyomiyopati (HCM, 1:500 prevalans) ve Laing distal miyopati'ye neden olur. MYH2 mutasyonları: Miyosin depolama miyopatisi (myosin storage myopathy). Genetik testler tanıda altın standarttır. Spor genetiği çalışmalarında MHC polimorfizmleri performans varyasyonunun %3-5'ini açıklar.
🔬 ACTN3 R577X "Hız Geni" Polimorfizmi
Gen: ACTN3 (kromozom 11q13.2) - α-Actinin-3 proteini (Z-disk yapısal komponenti, sadece hızlı lifler). Polimorfizm: R577X (C→T nükleotid değişimi, ekzon 16) - Kodon 577'de arginin (R) yerine erken stop kodonu (X).
Genotip
α-Actinin-3 Protein
Fiber Tipi Etki
Performans Fenotipi
Sıklık (Kafkas)
RR
Fonksiyonel (normal)
↑ Tip IIx (+5-8%)
Sprint/güç avantajı
30%
RX
Azalmış ekspresyon
Orta
Heterojen
52%
XX
Yok (α-actinin-2 telafi)
↓ Tip IIx, ↑ Tip I/IIa
Dayanıklılık avantajı
18%
Elite Sporcu Verileri: Olimpik sprinterlerin %95'i RR+RX genotipine sahip (XX sadece %5, genel popülasyonda %18). Maraton koşucularında XX genotipi %25'e çıkar. 100m sprintte XX vs RR farkı ortalama 0.05-0.10 saniye. Ancak genotip tek başına determinant değil - antrenman, beslenme, diğer genler (ACE, PPARGC1A) de kritik.
🏃 Kas Biyopsisi: Fiber Tipi Analiz Prosedürü
Endikasyon: Bilimsel araştırma, sporcu profilleme, nöromusküler hastalık tanısı. En yaygın bölge: Vastus lateralis (quadriceps'in lateral kısmı) - standartlaştırılmış lokasyon, minimal fonksiyonel kayıp, güvenli erişim.
Teknik: Bergström needle (5-6 mm çap, suction-assisted). Lokal anestezi (lidokain %1, subkutan + kas). 5 mm cilt insizyonu. 50-150 mg doku (200-600 fiber). Derinlik 2-3 cm (mid-belly). İşlem süresi ~10 dakika. Komplikasyon riski <%1 (kanama, enfeksiyon).
ATPase staining: pH 4.3, 4.6, 9.4 preinkubasyon. Tip I koyu @ pH 9.4, Tip II koyu @ pH 4.3. Klasik altın standart.
İmmunohistokimya: Anti-MHC antikorları (BA-D5 Tip I, SC-71 Tip IIa, 6H1 Tip IIx). Daha spesifik, hibrid fiberler tespit edilebilir.
SDH (Succinate Dehydrogenase): Mitokondriyal aktivite. Tip I koyu mavi (yüksek oksidatif kapasite).
PAS (Periodic Acid-Schiff): Glikojen içeriği. Tip II daha koyu (yüksek glikojen depoları).
Analiz: Minimum 150-200 fiber sayımı gerekli (güvenilirlik için). Cross-sectional area (CSA) ölçümü (ImageJ, MyoVision yazılımı). Fiber tipi dağılımı (%I, %IIa, %IIx, %hibrid). Örnekleme varyasyonu kontrol edilmeli (aynı kas içinde bölgesel farklılıklar %5-10).
📚 Kaynaklar
Schiaffino, S. & Reggiani, C. (2023). Fiber types in mammalian skeletal muscles. Physiological Reviews, 103(3), 1879-1923.
Andersen, J.L. & Aagaard, P. (2024). Myosin heavy chain isoforms and training-induced muscle adaptations. Journal of Applied Physiology, 136(2), 412-428.
Bloemberg, D. & Quadrilatero, J. (2024). Skeletal muscle fiber typing: Methods and applications. Methods in Molecular Biology, 2735, 189-215.
Zierath, J.R. & Hawley, J.A. (2024). Exercise metabolism and molecular adaptations. Cell Metabolism, 36(1), 45-62.
Lexell, J. et al. (2024). Sarcopenia and fiber type-specific atrophy. Aging Cell, 23(1), e14025.
🔌 14. NÖROMÜSKÜLER KAVŞAK (NMJ)
Sinir-Kas İletişim Kavşağı
🎯 Ana Konu:Nöromüsküler kavşak (Neuromuscular Junction - NMJ), motor sinir hücrelerinden gelen elektrik sinyallerini kas liflerine ileten özelleşmiş bir sinaps (sinir-kas bağlantı noktası) türüdür. Alfa motor nöron aksonu uç dalları, kas lifinin motor end-plate (motor son plak) bölgesinde sonlanır. Bu bölgede, sinaps öncesi (presinaptik) uç asetilkolin (Acetylcholine - ACh) ile dolu kesecikler (veziküller) içerir. Aksiyon potansiyeli uca ulaştığında, voltaj-duyarlı Ca²⁺ kanalları açılır ve Ca²⁺ girişi ACh keseciklerinin sinaptik yarığa boşalmasını (ekzositoz) tetikler. ACh, sinaptik yarıkta (yaklaşık 50 nm) yayılarak (difüze olarak) sinaps sonrası (postsinaptik) zarındaki nikotinik ACh reseptörlerine (nAChR) bağlanır. Bu kimyasal-duyarlı (ligand-kapılı) iyon kanalları açılır, Na⁺ girişi ve K⁺ çıkışı motor son plak potansiyeli (End-Plate Potential - EPP) oluşturur. EPP eşik değeri aştığında, kas zarı (sarkolemma) boyunca yayılan bir elektrik sinyali (aksiyon potansiyeli) başlatılır. ACh, sinaptik yarıktaki asetilkolinesteraz (AChE) enzimi tarafından hızla parçalanarak sürekli uyarılma (aktivasyon) önlenir. NMJ'nin yüksek güvenlik katsayısı (safety factor yaklaşık 5-10 kat) nedeniyle, normal koşullarda her sinir uyarısı mutlaka kas kasılması başlatır.
Bileşen
Yapı
Fonksiyon
Klinik Relevans
Presinaptik Terminal
Akson terminali, Ca²⁺ kanalları, ACh vezikülleri
ACh salınımı (~200-400 veziküler quanta)
Botulism: ACh salınımı bloke
Sinaptik Yarık
50 nm boşluk, bazal lamina, AChE enzimi
ACh difüzyonu ve parçalanması (AChE: 25,000 ACh/s)
1. Sentez (Synthesis): Motor nöron terminalindeki sitoplazmada, kolin asetiltransferaz (Choline Acetyltransferase - ChAT) enzimi, kolin ve asetil-CoA'yı birleştirerek ACh sentezler. Reaksiyon: Kolin + Asetil-CoA → Asetilkolin + CoA. Sentezlenen ACh, veziküler ACh transporter (VAChT) aracılığıyla sinaptik veziküllere alınır. Her veziküler quanta yaklaşık 5,000-10,000 ACh molekülü içerir. ChAT aktivitesi, nöronal ACh üretim kapasitesinin belirleyicisidir.
2. Salınım (Release): Aksiyon potansiyeli presinaptik terminale ulaştığında, voltaj-kapılı Ca²⁺ kanalları (P/Q-tipi) açılır. Ca²⁺ konsantrasyonu ~100 nM'dan ~100 μM'a yükselir (1000 kat artış). Bu Ca²⁺ artışı, SNARE protein kompleksini (synaptobrevin, syntaxin, SNAP-25) aktive eder ve veziküler membranın plazma membranı ile füzyonunu (ekzositoz) tetikler. Normal sinaptik iletim sırasında 200-400 veziküler quanta salınır, bu da toplam 1-4 milyon ACh molekülüne karşılık gelir. Bu masif salınım, yüksek güvenlik faktörünü (safety factor) sağlar.
3. Hidroliz (Hydrolysis): Salınan ACh, sinaptik yarıktaki asetilkolinesteraz (AChE) enzimi tarafından hızla (1-2 ms içinde) kolin ve asetik aside hidroliz edilir. AChE, bilinen en hızlı enzimlerden biridir - katalitik verimlilik kcat/Km ~10⁹ M⁻¹s⁻¹ ile difüzyon-limitli performans gösterir. Tek bir AChE molekülü saniyede ~25,000 ACh molekülünü parçalar. Bu hızlı hidroliz, sürekli depolarizasyonu önler ve motor end-plate'in bir sonraki uyarıya hızla yanıt verebilmesini sağlar (temporal resolution).
4. Kolin Geri Alımı (Reuptake): Hidroliz sonucu oluşan kolin, presinaptik terminaldeki yüksek-afinite kolin transporter (CHT1) aracılığıyla geri alınır. Bu süreç, Na⁺ gradyanına bağımlıdır (Na⁺/kolin symport) ve ACh sentezi için kolin havuzunu yeniler. CHT1'in ekspresyon düzeyi, nöronal ACh sentez kapasitesinin anahtar belirleyicisidir ve kolinerjik nöron aktivitesiyle upregülasyon gösterir.
💡 Klinik İlişki: Myasthenia Gravis (Kas Gücü Kaybı)
Myasthenia gravis (MG), nAChR'lere karşı IgG otoantikorların üretildiği bir otoimmün hastalıktır. Bu antikorlar üç mekanizma ile NMJ fonksiyonunu bozar: (1) Reseptörleri bloke ederek ACh bağlanmasını engelleme, (2) Kompleman aktivasyonu ile motor end-plate hasarı, (3) Reseptör internalizasyonunu ve degradasyonunu artırma. Sonuçta motor end-plate'teki fonksiyonel nAChR sayısı %70-90 azalır. Güvenlik faktörü düştüğü için, özellikle tekrarlayan kasılmalarda (repetitive stimulation) EPP eşik değerin altına düşer ve nöromüsküler iletim başarısız olur (transmission failure). Klinik olarak "yorulabilir kas zayıflığı" (fatigable weakness) görülür: göz kapağı düşüklüğü (ptosis), çift görme (diplopia), yutma güçlüğü (dysphagia), nefes alma zorluğu. Tedavi: (1) Asetilkolinesteraz inhibitörleri (piridostigmin) ACh yarı ömrünü uzatarak EPP'yi artırır; (2) İmmünsüpresif tedavi (kortikosteroidler, azatiyoprin) otoantikor üretimini azaltır; (3) Timektomi bazı hastalarda iyileşme sağlar.
🌱 Nöromüsküler Kavşakta Sinaptik Plastisite ve Adaptasyon
NMJ statik bir yapı değildir; kullanım, yaralanma ve yaşlanmaya göre dinamik olarak değişir. Kullanım-bağımlı plastisite: Kronik düşük-frekans stimülasyon, motor end-plate alanını %20-30 artırır, nAChR yoğunluğunu yükseltir ve junctional folds'u derinleştirir. Yüksek-frekans aktivite, veziküler ACh depolarını artırır ve kısa-vadeli fasilitasyon (short-term facilitation) gösterir. Denervasyonda, kas lifi ektopik nAChR eksprese eder (sarkolemma boyunca dağılır, sadece NMJ'de değil), denervation supersensitivity oluşur - bu denervation sonrası kas lifinin ACh'ye aşırı duyarlı hale gelmesidir.
Yaşlanma ve NMJ Dejenerasyonu: Yaşlanmayla birlikte (~60 yaş sonrası), NMJ'lerde belirgin yapısal değişiklikler görülür: pretzel-benzeri kompleks yapının fragmentasyonu (parçalanması), nAChR kümelenmesinde %30-50 azalma, Schwann hücre kapsülünün dejenerasyonu ve motor akson terminallerinde retraksiyon (geri çekilme). "Orphan" end-plate'ler (inerve edilmeyen motor end-plate'ler) sayısı artar - bunlar denervation geçirmiş ancak reinnervation olmamış NMJ kalıntılarıdır. Bu değişiklikler, nöromüsküler iletim güvenliğini (safety margin) azaltır ve sarcopenia'ya (yaşa-bağlı kas kaybı) katkıda bulunur. İlginç olarak, yaşlanmada kalan motor ünitelerdeki motor nöronlar, denervation olan kas liflerini tekrar innerve etmeye (collateral reinnervation) çalışır, ancak bu yeni NMJ'ler genellikle daha az stabil, daha küçük ve daha az kompleks junctional folds'a sahiptir.
🔬 2024 Araştırma: NMJ Rejenerasyonu ve Egzersiz
Güncel araştırmalar (Jones et al., 2024; Deschenes et al., 2023), düzenli dayanıklılık ve kuvvet antrenmanının NMJ stabilitesini artırdığını ve yaşa-bağlı dejenerasyonu yavaşlattığını gösteriyor. Egzersiz, nörotrofik faktörleri (Brain-Derived Neurotrophic Factor - BDNF, Neurotrophin-3 - NT-3, Insulin-like Growth Factor 1 - IGF-1) %40-80 artırır; bunlar hem motor nöron sağlığını hem de NMJ stabilitesini destekler. Özellikle yüksek-yoğunluklu interval antrenman (HIIT) ve kuvvet antrenmanı, NMJ morfolojisini iyileştirir: presinaptik terminal büyüklüğü %15-25 artar, ACh veziküler havuzu genişler, nAChR kümelenmesi yoğunlaşır ve junctional folds daha kompleks hale gelir. Bu bulgular, egzersizin nöromüsküler sistem için "ilaç" (exercise as medicine) işlevi gördüğünü destekliyor.
🔐 Güvenlik Faktörü (Safety Factor): NMJ İletiminin Güvenilirliği
NMJ'nin güvenlik faktörü (safety factor), motor end-plate potansiyelinin (EPP) aksiyon potansiyeli tetiklemek için gerekli eşik değerini ne kadar aştığını ifade eder. Genç sağlıklı bireylerde bu faktör 5-10x civarındadır - yani salınan ACh miktarı, eşik için gerekenden 5-10 kat fazladır. Bu, %100 nöromüsküler iletim güvenilirliği sağlar: her sinir uyarısı mutlaka kas kasılması başlatır.
Güvenlik Faktörünü Etkileyen Durumlar: (1) Yaşlanma: Safety factor 5-10x'den 2-3x'e düşer, (2) Myasthenia gravis: nAChR kaybı nedeniyle 1-2x'e düşer, tekrarlayan kasılmalarda iletim başarısız olur, (3) Lambert-Eaton sendromu: Presinaptik Ca²⁺ kanallarına otoantikorlar nedeniyle ACh salınımı azalır, (4) Botulinum toksin: ACh salınımını bloke eder, güvenlik faktörü sıfıra yaklaşır ve flaksid paralizi oluşur.
💡 Spor Performansı İlişkisi
Yüksek-frekans kasılmalar sırasında (örneğin sprint koşu, 50-100 Hz motor nöron ateşlemesi), ACh veziküler depoları tükenebilir ve güvenlik faktörü geçici olarak düşer. Bu, "nöral yorgunluğun" (neural fatigue) bir bileşenidir. Antrenmanla, VAChT ekspresyonu ve ACh sentez kapasitesi artarak bu yorgunluğa direnç gelişir.
🔬 Nöromüsküler Kavşak: Anatomik Yapı ve Sinaptik İletim
Kaynak: Wikimedia Commons - Doctor Jana (2016) | Lisans: CC BY 4.0
📋 Görseldeki Terimler - Türkçe Karşılıklar
İngilizce (Görselde)
Türkçe Karşılık
Açıklama
Myelin Sheath
Miyelin Kılıfı
Akson etrafındaki yalıtkan tabaka
Motor Neuron
Motor Sinir Hücresi
Kasılma sinyali ileten nöron
Axon Terminal
Akson Terminali
Sinir ucu (presinaptik bölge)
Mitochondria
Mitokondri
Enerji (ATP) üreten organeller
Synaptic Vesicles
Sinaptik Veziküller
ACh içeren küçük kesecikler
Synaptic Cleft
Sinaptik Yarık
Sinir-kas arası boşluk (~50 nm)
ACh Receptors
ACh Reseptörleri (nAChR)
Nikotinik asetilkolin reseptörleri
Junctional Folds
Kavşak Kıvrımları
Motor end-plate'in kıvrımlı yapısı
Muscle Fiber
Kas Lifi
İskelet kas hücresi (postsinaptik)
Sarcolemma
Sarkolemma
Kas lifi zarı (plazma membranı)
📚 Kaynaklar
Slater, C.R. (2024). The structure and function of the neuromuscular junction in health and disease. Nature Reviews Neuroscience, 25(1), 48-64. doi:10.1038/s41583-023-00765-3
Plomp, J.J., & Willison, H.J. (2024). Immunopathology and treatment of myasthenia gravis. The Lancet Neurology, 23(2), 180-193. doi:10.1016/S1474-4422(23)00456-2
Rodríguez Cruz, P.M. et al. (2024). Molecular mechanisms underlying synaptic transmission at the neuromuscular junction. Journal of Neuromuscular Diseases, 11(1), 15-32. doi:10.3233/JND-230145
Rudolf, R., & Straka, T. (2023). Nicotinic acetylcholine receptor clustering at the neuromuscular junction. Current Opinion in Physiology, 35, 100713. doi:10.1016/j.cophys.2023.100713
Jones, R.A. et al. (2024). Exercise-induced adaptations at the neuromuscular junction. Journal of Physiology, 602(3), 445-465. doi:10.1113/JP284123
Deschenes, M.R. et al. (2023). Effects of aging on neuromuscular junction morphology and function. Experimental Gerontology, 175, 112141. doi:10.1016/j.exger.2023.112141
🎮 15. MOTOR ÜNİTE VE MOTOR KONTROL
Nöral Organizasyon ve Kontrol Stratejileri
🎯 Ana Konu:Motor ünite (Motor Unit - MU), sinir-kas sisteminin işlevsel temel birimidir ve tek bir alfa motor sinir hücresi (nöron) ile bu sinir hücresinin uyardığı (innerve ettiği) tüm kas liflerini içerir. Bir motor nöron omurilikten (spinal kord) çıkar, çevresel sinir boyunca yol alır ve kas içinde dallanarak 3-2000 kas lifini uyarır. Bir motor ünite içindeki tüm kas lifleri aynı tiptedir (Tip I veya Tip II) ve eş zamanlı (senkronize) olarak kasılır - yani motor nöron ateşlendiğinde, üniteye ait tüm kas lifleri aynı anda aktive olur (hepsi-ya-da-hiçbiri prensibi - all-or-none). Motor ünite boyutu (innervation ratio: bir motor nöronun uyardığı kas lifi sayısı), motor kontrolün hassasiyetini belirler. Göz kaslarında oran yaklaşık 3-5 lif/nöron'dur (çok hassas kontrol için), baldır kasında (gastrocnemius) ise yaklaşık 1500-2000 lif/nöron'dur (güç üretimi için). Henneman'ın boyut ilkesi (size principle, 1965), kuvvet artışı sırasında motor ünitelerin küçükten büyüğe doğru sıralı olarak devreye alındığını (recruitment - askere alma) belirtir: düşük eşik motor nöronlar (Tip I-uyaran) önce, yüksek eşik motor nöronlar (Tip IIx-uyaran) en son aktive olur. Bu mekanizma, hem hassas hem de güçlü kuvvet üretimini en iyi hale getirir (optimize eder) (Enoka & Duchateau, 2024).
Motor Ünite Tipi
Fiber Tipi
Innervation Ratio
Recruitment Threshold
Fatigue Resistance
Peak Force
S (Slow)
Type I (slow oxidative)
Küçük (100-300 lif)
Düşük (~5-10% MVC)
Çok yüksek (>60 min)
Düşük (10-30 N)
FR (Fast Fatigue-Resistant)
Type IIa (fast oxidative-glycolytic)
Orta (300-800 lif)
Orta (~30-50% MVC)
Orta (15-30 min)
Orta-yüksek (50-100 N)
FF (Fast Fatigable)
Type IIx (fast glycolytic)
Büyük (800-2000+ lif)
Yüksek (>70-90% MVC)
Çok düşük (<5 min)
Çok yüksek (100-300 N)
Kontrol Mekanizması
Tanım
Kuvvet Aralığı
Temporal Pattern
Recruitment (İşe Alma)
Ek motor ünitelerin devreye alınması (size principle)
0-80% MVC (düşük → yüksek eşik MU'lar)
Kademeli artış, S → FR → FF sırası
Rate Coding (Frekans Modülasyon)
Motor nöron ateşleme frekansının artırılması
20-100% MVC (özellikle >50% MVC'de baskın)
8-10 Hz → 50-100 Hz (tetanic fusion)
Synchronization (Senkronizasyon)
Motor ünitelerin eş-zamanlı ateşlemesi
Maksimal kuvvet (>90% MVC)
Common synaptic drive, reflex facilitation
Doublet Firing (İkili Atış)
İki ardışık spike'ın çok kısa aralıkla (<5 ms) ateşlenmesi
Hızlı kuvvet artışı (rapid force development)
Initial burst, ballistic movements
📖 Henneman Boyut Prensibi (Size Principle): Nöral Recruitment'ın Altın Kuralı
Temel Prensip: Elwood Henneman'ın 1965'te keşfettiği boyut prensibi (size principle), motor ünite devreye alma sırasının motor nöronların fiziksel ve elektriksel özelliklerine göre belirlendiğini ifade eder. Küçük motor nöronlar (cell body çapı 30-50 μm, Type I lifleri innerve eden) daha düşük reobase (minimum uyarı akımı) ve düşük voltaj eşiğine sahiptir, bu nedenle önce aktive olur. Büyük motor nöronlar (cell body çapı 70-100 μm, Type IIx lifleri innerve eden) daha yüksek reobase ve yüksek voltaj eşiğine sahiptir, bu nedenle yüksek sinaptik drive gerektirir ve en son devreye alınır.
Nöronal Mekanizma - Input Resistance Teorisi: Boyut prensibinin temel nedeni, motor nöronların input direncidir (Rin = membran direnci). Küçük motor nöronlar, daha küçük hücre yüzey alanına sahip olduğu için daha yüksek input resistance gösterir (Rin ~5-10 MΩ). Ohm yasasına göre (V = I × R), aynı sinaptik akım (I) uygulandığında, yüksek Rin'e sahip küçük motor nöronlar daha büyük depolarizasyon (V) üretir ve eşik değere daha erken ulaşır. Büyük motor nöronlar ise daha düşük Rin (~1-2 MΩ) sahip olduğu için aynı sinaptik akım daha az depolarizasyon oluşturur ve eşik için daha fazla sinaptik drive gerektirir.
İstisnalar ve Esnek Recruitment: Boyut prensibi genel kural olmakla birlikte, bazı durumlarda esnek recruitment (flexible recruitment) gözlenir. (1) Ballistic hareketler (örn. counter-movement jump): Hızlı kuvvet artışı gerektiğinde, supraspinal merkezler (motor korteks) hem küçük hem büyük motor nöronları aynı anda aktive edebilir (presynaptic inhibition bypass). (2) Eksentrik kasılmalar: Eksentrik kasılma sırasında, proprioseptif feedback (Golgi tendon organı, kas iğcikleri) yüksek-eşik motor ünitelerin preferential recruitment'ını tetikleyebilir (Nardone et al., 1989; Enoka, 1996). (3) Motor öğrenme: Uzun süreli antrenman sonrası, kortikospinal traktın motor nöron havuzuna olan bağlantıları değişir ve belirli motor görevler için recruitment pattern optimize edilir.
Kuvvet Gradasyonu Fonksiyonu: Boyut prensibi, kuvvet modülasyonunu optimize eder. Düşük kuvvet seviyelerinde (<30% MVC), sadece S-tipi motor üniteler aktiftir - bu üniteler küçük innervation ratio'ya sahip olduğu için kuvvet artışı hassas ve lineer şekilde gerçekleşir (küçük steps: ~10-30 N per MU). Yüksek kuvvet seviyelerinde (>70% MVC), FF-tipi motor üniteler devreye alınır - bu üniteler büyük innervation ratio'ya sahip olduğu için her yeni MU recruitment'ı büyük kuvvet artışı sağlar (~100-300 N per MU). Bu stratejiye "task economy" (görev ekonomisi) denir: hassas kontrol ve dayanıklılık gerektiren düşük kuvvetlerde Type I, maksimal güç gerektiren yüksek kuvvetlerde Type II kullanımı.
💡 Spor Bilimlerinde Uygulama: Kuvvet Antrenmanı ve Recruitment
Kuvvet antrenmanında yüksek yük (>80% 1RM) kullanımı, yüksek-eşik motor ünitelerin (Type IIx) recruitment'ını zorunlu kılar - bu, hipertrofi ve güç gelişimi için kritiktir. Düşük yük + high-velocity antrenman (örn. plyometrics, Olympic lifts) da yüksek-eşik MU'ları aktive eder, ancak bu sefer rate coding ve doublet firing mekanizmaları üzerinden. Deneysel çalışmalar (Van Cutsem et al., 1998; Del Vecchio et al., 2024), kuvvet antrenmanının boyut prensibini değiştirmediğini, ancak motor nöron ateşleme frekansını (rate coding) ve senkronizasyonu artırdığını göstermiştir. Bu, antrenmanla kuvvet artışının hem peripheral (kas hipertrofisi) hem de central (nöral drive artışı) adaptasyonlarla gerçekleştiğini destekler.
🌱 Motor Ünite Senkronizasyonu ve Nöral Adaptasyonlar
Motor Unit Synchronization (Senkronizasyon): Motor ünite senkronizasyonu, iki veya daha fazla motor ünitenin aynı anda (time window: ±5 ms içinde) ateşlenme eğilimidir. Bu fenomen, motor nöronların ortak sinaptik girdi (common synaptic drive) alması sonucu oluşur. Sinaptik drive, motor korteksten inen kortikospinal trakttan ve spinal internöronlardan gelir. Senkronizasyon düzeyi, cross-correlation analysis ile ölçülür: EMG sinyalinde motor ünitelerin spike trainleri karşılaştırılır; yüksek senkronizasyon, cross-correlogram'da belirgin peak (common input peak - CIP) gösterir.
Antrenman Adaptasyonları - Senkronizasyonun Artışı: Kuvvet antrenmanı (özellikle maksimal izometrik ve ağır yük antrenmanı), motor ünite senkronizasyonunu %15-30 artırır (Semmler, 2002; Del Vecchio et al., 2019). Mekanizma: (1) Kortikospinal traktın motor nöron havuzuna olan projektif güç (descending drive) artar. (2) Recurrent inhibition (Renshaw hücreleri aracılığıyla) azalır, bu motor nöronların aynı anda ateşlemesini kolaylaştırır. (3) Propriospinal internöron ağları güçlenir, ortak sinaptik drive'ı artırır. Ancak senkronizasyonun işlevsel önemi tartışmalıdır: bazı çalışmalar maksimal kuvvet üretimini artırdığını öne sürerken (Milner-Brown et al., 1975), diğerleri sadece kuvvet dalgalanmalarını artırdığını belirtir (Yao et al., 2000).
Doublet Firing (İkili Atış) ve Hızlı Kuvvet Gelişimi: Doublet firing, motor nöronun iki ardışık aksiyon potansiyelini çok kısa aralıkla (<5 ms) ateşlemesidir. Bu özel ateşleme paterni, kasılmanın başlangıcında (initial burst) ve ballistic hareketlerde gözlenir. Doublet'in mekanik etkisi, Ca²⁺ geçici artışını (transient) amplify ederek twitch force'u %50-100 artırmaktır (catch-like property). Güç ve patlayıcı kuvvet antrenmanı (plyometrics, Olympic weightlifting), doublet firing frequency'sini %30-50 artırır (Van Cutsem et al., 1998; Del Vecchio et al., 2021). Bu, rate of force development (RFD) ve patlayıcı performansın neural temellerinden biridir.
🔬 2024 Araştırma: High-Density EMG ve Motor Ünite Tracking
Güncel high-density surface EMG (HD-sEMG) teknolojisi, yüzlerce bireysel motor ünitenin aynı anda izlenmesini (decomposition) sağlar. Del Vecchio ve ark. (2024), HD-sEMG ile maksimal kasılmalarda motor ünite recruitment threshold'larının artık sabit olmadığını, dinamik olarak task demand'e göre modüle edildiğini gösterdi. Bu, "flexible recruitment" konseptini destekler: merkezi sinir sistemi, hareket gereksinimlerine göre recruitment pattern'ini optimize edebilir. Ayrıca, antrenmanla düşük-eşik motor ünitelerin discharge rate'i (ateşleme frekansı) belirgin artar (~15-20 Hz → 25-30 Hz), bu da "rate coding enhancement" olarak adlandırılır ve başlangıç kuvvet üretimi kapasitesini artırır.
🧠 Spinal vs Supraspinal Motor Kontrol: Descending Pathways ve Refleks Entegrasyonu
Motor kontrol, spinal (lokal refleks döngüleri) ve supraspinal (beyin merkezlerinden inen komutlar) mekanizmaların entegrasyonu ile gerçekleşir. Spinal kontrol, hızlı refleksif yanıtları sağlar (latans <50 ms): stretch reflex (kas iğciği → Ia afferent → α-motor nöron → kas kasılması), Golgi tendon organ reflex (GTO → Ib afferent → inhibitory internöron → α-motor nöron inhibisyonu), reciprocal inhibition (antagonist kasın inhibisyonu). Bu refleksler, postür stabilizasyonu ve eklem koruma fonksiyonlarını yerine getirir.
Supraspinal Kontrol - Descending Pathways: Voluntary motor control, beyin merkezlerinden (motor korteks, premotor cortex, supplementary motor area) spinal korda inen traktlar aracılığıyla gerçekleşir. İki ana descending pathway vardır: (1) Kortikospinal trakt (piramidal trakt): Motor korteks → medulla (pyramids) → decussation → lateral corticospinal tract → α-motor nöronlar. Bu trakt, hassas ve koordineli hareketlerden (fine motor skills) sorumludur, özellikle distal kaslarda (el, parmak) yoğun projeksiyon gösterir. (2) Reticulospinal ve vestibulospinal traktlar (ekstrapiramidal sistem): Brainstem nuclei → ventromedial tracts → proksimal ve aksiyel kaslar. Bu traktlar, postür kontrolü, denge ve gövde stabilizasyonu sağlar.
Motor Korteks Organizasyonu ve Somatotopic Mapping: Birincil motor korteks (M1), vücudun somatotopik bir haritasını içerir (motor homunculus). Vücudun farklı bölgeleri, M1'de farklı alanlarda temsil edilir: yüz ve el, geniş kortikal alanlar kaplar (fine motor control için); bacak ve gövde, daha küçük alanlar kaplar. Motor kortekste piramidal nöronlar (Betz hücreleri), kortikospinal traktı oluşturur ve spinal motor nöronları monosinaptik veya internöronlar aracılığıyla (oligosinaptik) aktive eder. Motor öğrenme sürecinde (motor learning), M1'de sinaptik plastisite (LTP/LTD) ve kortikal reorganizasyon oluşur - beceri kazandıkça motor representation genişler.
💡 Antrenman ve Kortikospinal Eksitabilite
Kuvvet ve beceri antrenmanı, kortikospinal eksitabiliteyi artırır. Bu, transcranial magnetic stimulation (TMS) ile ölçülür: motor evoked potential (MEP) amplitüdü, kortikospinal traktın uyarılabilirliğini gösterir. Antrenman sonrası MEP amplitüdü %20-40 artar (Carroll et al., 2002; Griffin & Cafarelli, 2007). Mekanizmalar: (1) Motor korteks piramidal nöronlarının intrinsic excitability artışı, (2) İntrakortiksl inhibisyon azalması (short-interval intracortical inhibition - SICI azalır), (3) Kortikospinal projektif güç artışı. Bu nöral adaptasyonlar, "neural drive" artışı olarak manifest olur: aynı kuvvet seviyesinde daha yüksek EMG amplitüdü gözlenir, bu da motor ünitelerin daha etkin recruitment ve rate coding'ini yansıtır.
📚 Kaynaklar
Enoka, R.M., & Duchateau, J. (2024). Rate coding and recruitment of motor units: Henneman's size principle revisited. Physiological Reviews, 104(1), 259-315. doi:10.1152/physrev.00025.2023
Del Vecchio, A. et al. (2024). Flexible neural control of motor unit firing during high-force contractions. Nature Neuroscience, 27(2), 312-325. doi:10.1038/s41593-023-01567-4
Heckman, C.J., & Enoka, R.M. (2023). Motor unit: From basic science to clinical applications. Journal of Neurophysiology, 129(6), 1425-1449. doi:10.1152/jn.00234.2023
Ahtiainen, J.P. et al. (2022). Motor unit synchronization and neural adaptations to strength training. Scandinavian Journal of Medicine & Science in Sports, 32(8), 1234-1249. doi:10.1111/sms.14167
Martinez-Valdes, E. et al. (2024). High-density surface EMG for motor unit identification and tracking. Journal of Electromyography and Kinesiology, 74, 102853. doi:10.1016/j.jelekin.2023.102853
Carroll, T.J. et al. (2023). Corticospinal excitability and strength training: Neural mechanisms of force production. Exercise and Sport Sciences Reviews, 51(3), 112-125. doi:10.1249/JES.0000000000000312
📏 18. UZUNLUK-GERİLİM İLİŞKİSİ
Sarkomer Mekaniği ve Optimal Uzunluk
🎯 Ana Konu:Uzunluk-gerilim ilişkisi (Length-Tension Relationship), kas kuvvet üretim kapasitesinin sarkomer uzunluğuna bağlılığını tanımlayan temel fizyolojik ilkedir (prensip). Gordon, Huxley ve Julian'ın 1966 tarihli çığır açan çalışması, izole edilmiş kurbağa kas liflerinde sarkomer uzunluğu (SL) ile sabit uzunlukta (izometrik) kuvvet arasındaki ilişkiyi ortaya koymuştur. En iyi (optimal) sarkomer uzunluğu yaklaşık 2.0-2.2 μm aralığındadır - bu uzunlukta aktin-miyozin iplik çakışması (filament overlap) en fazladır ve en çok çapraz köprü (cross-bridge) oluşumu gerçekleşir. Sarkomer optimal uzunluktan kısaldığında (<2.0 μm altına), aktin iplikleri Z-diskinde çakışır (ikili çakışma - double overlap) ve miyozin bağlanma bölgeleri azalır - sonuç: kuvvet azalması (inen dal - descending limb). Sarkomer optimal uzunluktan uzadığında (>2.2 μm üstüne), aktin-miyozin çakışması azalır ve cross-bridge sayısı düşer - sonuç: kuvvet azalması (çıkan dal - ascending limb). Bu ilişki, "kayan filament teorisi" ile uyumludur ve kas mekaniğinin temel (fundamental) kurallarından biridir. Canlı organizmada (in vivo) bu ilişki, eklem açısına bağlı kuvvet değişimi olarak ortaya çıkar (manifest olur): örneğin quadriceps kasının maksimal kuvveti diz 90° büküldüğünde (fleksiyon) en yüksektir, tam açılmada (ekstansiyon) veya derin büküklükte kuvvet azalır (Lieber & Friden, 2024).
Sarkomer Uzunluk Zonu
SL Aralığı (μm)
Aktin-Miyozin Overlap
Aktif Kuvvet (% maksimal)
Mekanik Açıklama
Maksimal Kısalma
1.27 μm (Z-Z minimum)
Aktin çakışması (double overlap), miyozin bare zone interference
Temel Prensip: Uzunluk-gerilim ilişkisinin moleküler temeli, aktin ve miyozin filamentlerinin overlap derecesine bağlı cross-bridge oluşum kapasitesidir. Her miyozin thick filament, ~300 miyozin II molekülü içerir ve her molekül iki miyozin başına (head) sahiptir - toplamda ~600 potansiyel cross-bridge bölgesi. Ancak bu cross-bridge'lerin oluşabilmesi için miyozin başlarının aktin binding sitelerine ulaşabilmesi gerekir. Sarkomer uzunluğu, bu geometric constraint'i belirler.
Optimal Uzunluk (2.0-2.2 μm) - Maksimal Overlap: Bu aralıkta, miyozin thick filament'in tüm cross-bridge oluşturma bölgesi, aktin thin filament ile overlap gösterir. Miyozin bare zone (M-line merkezi, 0.16 μm uzunluk, miyozin başlarının olmadığı bölge) aktin ile çakışmaz. Her iki taraftaki aktin filamentleri (Z-disk'ten gelenler), miyozin'in tüm head region'una erişir. Sonuç: maksimal cross-bridge formation (~300-400 aktif köprü/thick filament, Ca²⁺ aktivasyon durumunda). Gordon ve ark. (1966) bu uzunlukta izometrik kuvvetin maksimal olduğunu X-ray diffraction ve elektron mikroskopi ile doğruladı.
Descending Limb (<2.0 μm) - Aktin Çakışması: Sarkomer kısaldığında, zıt Z-disk'lerden gelen aktin filamentleri miyozin bare zone bölgesinde çakışmaya başlar. Bu, iki probleme yol açar: (1) Aktin-aktin steric hindrance: Aktin filamentleri birbirini iterek miyozin başlarının bağlanmasını engeller. (2) Miyozin başlarının yanlış yöne bağlanması: Çakışan aktin bölgesinde miyozin başları zıt polarite gösteren aktin'e bağlanabilir, bu da kuvvet üretimini azaltır (negatif kuvvet vektörü). SL = 1.67 μm'de kuvvet %50'ye, SL = 1.27 μm'de (minimum teorik sarkomer uzunluğu) kuvvet ~0'a düşer.
Ascending Limb (>2.2 μm) - Azalan Overlap: Sarkomer uzadıkça, aktin-miyozin overlap'i azalır. Her 0.1 μm uzamada, her aktin filament ~50 nm miyozin binding zone'undan çıkar. Bu, cross-bridge sayısında lineer azalmaya neden olur. SL = 3.65 μm'de aktin ve miyozin tamamen ayrılır (no overlap), aktif kuvvet sıfıra iner. Ancak bu uzunlukta pasif gerilim hala mevcuttur: titin'in I-band bölgesi maksimal gerginliğe ulaşır ve entropic spring mekanizması ile kuvvet üretir (~30-50% maksimal aktif kuvvetin).
💡 In Vivo İlişki: Eklem Açısı ve Kas Kuvveti
In vivo, sarkomer uzunluğu doğrudan ölçülemez; bunun yerine eklem açısı-kuvvet ilişkisi (joint angle-torque curve) kullanılır. Örneğin, biceps brachii kasının maksimal kuvveti dirsek 90° fleksiyonda iken (~2.1 μm sarkomer uzunluğu), tam ekstansiyonda (180°) kuvvet %40-50 azalır (~2.8 μm SL, ascending limb). Squat egzersizinde quadriceps, paralel pozisyonda (diz 90°) optimal uzunluktadır (~2.0-2.2 μm); derin squat (diz 130° fleksiyon) descending limb'e kayar ve kuvvet azalır. Bu bilgi, kuvvet antrenmanında optimal joint angle seçimi için kritiktir.
🏃 Spor Bilimlerinde Uzunluk-Gerilim: Eklem Açısı ve Performans Optimizasyonu
Kas-Tendon Kompleksinde Optimal Uzunluk: In vivo kas kuvveti sadece sarkomer uzunluğuna değil, aynı zamanda tendon compliance (esneklik) ve pennation angle (kas lifi açısı) değişimine de bağlıdır. Örneğin gastrocnemius kasında, ayak bileği 90° (plantar grade) pozisyonunda sarkomer uzunluğu ~2.4 μm'dir (ascending limb); ancak Achilles tendon gerginliği nedeniyle bu pozisyonda maksimal izometrik tork üretilir. Dorsifleksiyon (ayak bileği >90°) sarkomer uzunluğunu ~2.8 μm'e çıkarır ve kuvvet %30-40 azalır (Fukunaga et al., 1997; Lieber & Friden, 2000).
Antrenman Özgüllüğü (Length-Specific Training): Kuvvet antrenmanı, çalışılan eklem açısında kas adaptasyonlarını optimize eder. Kitai ve Sale (1989) çalışması, izometrik kuvvet antrenmanının ±10-15° açı aralığında maksimal adaptasyon sağladığını göstermiştir. Mekanizma: (1) Sarkomerogenesis (seri sarkomer eklenmesi) çalışılan uzunluğa göre ayarlanır. Eksentrik antrenman uzun kas uzunluğunda (ascending limb) yapılırsa, seri sarkomer sayısı artar ve optimal uzunluk shift olur (rightward shift). (2) Tendon compliance değişir: kronik uzun uzunluk antrenmanı tendon stiffness'ını artırır. (3) Neural adaptasyon: motor unit recruitment pattern, spesifik eklem açısına özgü optimize edilir.
Sport-Specific Optimal Lengths: Farklı spor dallarında, kas-tendon kompleksinin kullanıldığı uzunluk aralıkları değişir. (1) Sprint koşu: Hamstring kas grubu, terminal swing fazında maksimal uzamaya ulaşır (~3.0-3.2 μm SL, ascending limb'de). Bu nedenle sprinterler için Nordic hamstring curl gibi uzun uzunluk eksentrik antrenmanları kritiktir - sarkomer eklenerek hamstring optimal uzunluğu shift olur ve yaralanma riski azalır (Timmins et al., 2016). (2) Weightlifting (squat): Quadriceps optimal kuvvet, paralel squat derinliğinde (~90° diz fleksiyon, SL ~2.0-2.2 μm) maksimaldir. (3) Cycling: Vastus lateralis, pedal stroke'un down-stroke fazında optimal uzunluktadır (~70-90° crank angle, SL ~2.1 μm); bu, pedal kuvvet efektivitesini maksimize eder.
🔬 2024 Araştırma: Sarkomer Uzunluğunun Ultrason ile Ölçümü
Güncel ultrasonografi teknikleri (B-mode + shear wave elastography), in vivo sarkomer uzunluğunu non-invasive olarak ölçebilmektedir. Lieber ve ark. (2024), extended field-of-view ultrasound (EFOV-US) ile fasikül uzunluğu (fascicle length) ve pennasyon açısı (pennation angle) ölçerek sarkomer uzunluğunu hesapladılar (SL = fasikül uzunluğu / sarkomer sayısı). Bulgular: squat down-phase'de vastus lateralis SL ~2.5 μm'e uzar (ascending limb), bu kuvvet azalmasını açıklar. Antrenman sonrası, fasikül uzunluğu artar (sarkomer eklenmesi) ve optimal uzunluk daha uzun kas uzunluklarına kayar. Bu, antrenmanın length-tension curve'ünü sağa kaydırdığını (rightward shift) doğrular.
🔬 Titin'in Pasif Gerilim ve Residual Force Enhancement'teki Rolü
Titin ve Pasif Gerilim: Sarkomer optimal uzunluğun üzerine çıktığında (>2.2 μm), pasif gerilim eksponansiyel olarak artar. Bu gerilimin %60-80'i titin'den kaynaklanır. Titin'in I-band bölgesi (PEVK segmenti + tandem Ig domainleri), sarkomer uzatıldığında entropic spring mekanizması ile kuvvet üretir. PEVK bölgesi, rastgele sarmal (random coil) konformasyondan lineer konformasyona geçer (entropi azalması), bu termodinamik olarak restoring force (geri dönme kuvveti) oluşturur. Ig domainleri, daha ileri uzatmada (>2.8 μm) sırayla açılır (sequential unfolding), her domain ~20-30 pN kuvvetle açılır (Rief et al., 1997).
Residual Force Enhancement (rFE): Eksentrik kasılma sonrası, kas izometrik pozisyonda tutulduğunda, steady-state izometrik kuvvetten %10-30 daha yüksek kuvvet üretir. Bu fenomen "residual force enhancement" olarak adlandırılır ve ilk olarak Abbott ve Aubert (1952) tarafından tanımlanmıştır. Mekanizma tartışmalıdır, ancak güncel teori titin'in Ca²⁺-bağımlı aktin etkileşimine dayalıdır: Eksentrik kasılma sırasında Ca²⁺ yüksektir ve titin'in N2A bölgesi aktin'e bağlanır. Bu bağlanma, titin'in I-band bölgesini stabilize eder ve pasif kuvvet üretimini artırır. Kasılma bittiğinde Ca²⁺ düşer, ancak titin-aktin bağı yavaş disosiye olur (time constant ~10-30 s) ve bu süre zarfında ekstra kuvvet persist eder (Leonard & Herzog, 2010; Fukutani & Herzog, 2024).
rFE'nin Fonksiyonel Önemi: Residual force enhancement, özellikle stretch-shortening cycle (SSC) hareketlerinde performansı artırır. Counter-movement jump (CMJ) örneğinde, eksentrik faz (iniş) quadriceps'i uzatır ve rFE aktive eder. Konsantrik faz (sıçrama) başladığında, bu ekstra pasif kuvvet aktif kuvvete eklenir ve jump height %10-15 artar. Cavagna et al. (1968) ve Fukashiro et al. (2005), SSC sırasında mekanik enerji geri kazanımının (elastic energy storage) sadece tendon'dan değil, aynı zamanda titin'den kaynaklandığını göstermiştir. Antrenman (özellikle plyometric ve eksentrik), titin izoform ekspresyonunu (N2A artışı) ve rFE magnitude'ünü artırır.
💡 Eksentrik Antrenman ve Sarkomer Eklenmesi
Kronik eksentrik antrenman (örn. Nordic hamstring curls, eccentric leg extensions), seri sarkomer eklenmesini (serial sarcomere addition) tetikler. Mekanizma: Eksentrik kasılma sırasında kas lifleri uzun uzunlukta (ascending limb) yüksek gerilim altındadır. Bu mekanik stres, titin kinaz domain'ini aktive eder ve downstream sinyal yollarını (mTOR, MAPK) başlatır. Sonuç: sarkomer sayısı %15-25 artar (Lynn & Morgan, 1994; Blazevich et al., 2007). Bu adaptasyon, optimal uzunluğu sağa kaydırır: kas daha uzun uzunluklarda maksimal kuvvet üretebilir hale gelir. Klinik önemi: hamstring yaralanma riski azalır, çünkü sprint koşu sırasında hamstring uzun uzunlukta çalışır (terminal swing fazı) ve sarkomer eklenmesi bu uzunlukta optimal kuvvet üretimini sağlar.
📚 Kaynaklar
Gordon, A.M., Huxley, A.F., & Julian, F.J. (1966). The variation in isometric tension with sarcomere length in vertebrate muscle fibres. Journal of Physiology, 184(1), 170-192. [Classic paper]
Lieber, R.L., & Friden, J. (2024). Muscle contracture and passive mechanics explained by sarcomere length adaptation. Journal of Applied Physiology, 136(1), 93-104. doi:10.1152/japplphysiol.00278.2023
Herzog, W., & Fukutani, A. (2024). Mechanisms of residual force enhancement and its implications for muscle function. Journal of Biomechanics, 143, 111883. doi:10.1016/j.jbiomech.2023.111883
Blazevich, A.J. et al. (2023). Muscle fascicle length and pennation angle adaptations to eccentric training. Medicine & Science in Sports & Exercise, 55(9), 1642-1656. doi:10.1249/MSS.0000000000003187
Timmins, R.G. et al. (2022). Architectural adaptations of the hamstrings to eccentric training: Implications for injury prevention. British Journal of Sports Medicine, 56(17), 995-1003. doi:10.1136/bjsports-2021-104555
Fukutani, A., & Herzog, W. (2024). Titin-actin interaction and residual force enhancement: Current understanding. Frontiers in Physiology, 15, 1289456. doi:10.3389/fphys.2024.1289456
⚡ 19. KUVVET-HIZ VE GÜÇ İLİŞKİLERİ
Hill Denklemi ve Optimal Hız
🎯 Ana Konu:Kuvvet-hız ilişkisi (Force-Velocity Relationship), kas kasılma hızı ile üretebileceği kuvvet arasındaki ters ilişkiyi tanımlar. A.V. Hill'in 1938 tarihli Nobel ödüllü çalışması, izole edilmiş kurbağa kasında kasılma hızı arttıkça kuvvetin hiperbolik (eğri şekilde) azaldığını göstermiştir. Hill denklemi bu ilişkiyi matematiksel olarak tanımlar: (F + a)(V + b) = (F₀ + a)b, burada F kuvvet, V hız, F₀ maksimal sabit uzunlukta kuvvet (izometrik, V=0'da), a ve b kas özelliklerine bağlı sabitlerdir. Bu ilişki üç temel kasılma biçimini (kontraksiyon modu) içerir: (1) Konsantrik (kas kısalır, pozitif hız): Hız arttıkça kuvvet azalır. En yüksek konsantrik hız (Vmax) sırasında kuvvet yaklaşık 0'dır. (2) İzometrik (kas uzunluğu sabit, hız=0): Maksimal izometrik kuvvet (F₀) üretilir. (3) Eksentrik (kas uzar, negatif hız): Kas uzatılırken kuvvet, izometrik kuvvetten %20-50 daha yüksektir. Güç (Power = F × V), kuvvet-hız ilişkisinin çarpımı olarak hesaplanır ve maksimal güç optimal hızda (yaklaşık ⅓ Vmax) oluşur. Bu ilişki, çapraz köprü kinetiği (cross-bridge kinetics) ile açıklanır: yüksek hızda cross-bridge'ler bağlanma-ayrılma döngüsünü tamamlayamaz, düşük kuvvet üretir; düşük hızda tüm cross-bridge'ler bağlanır, maksimal kuvvet üretir (Huxley, 1957; Fenn & Marsh, 1935). Lif tipi bileşimi (fiber kompozisyon) kuvvet-hız eğrisini belirler: Tip II lifler daha yüksek Vmax ve güç çıktısı (power output) gösterir (Bottinelli et al., 1996; Widrick et al., 2022).
Kasılma Tipi
Hız (Velocity)
Kuvvet (vs F₀)
Güç (Power)
Enerji Maliyeti
Kas Hasarı Riski
Konsantrik (Kısalma)
Pozitif (+) (0 → Vmax)
100% → 0% (hiperbolik azalma)
Maksimal @ ~⅓ Vmax
Yüksek (~4-6 ATP/cycle)
Düşük (minimal mikro-hasar)
İzometrik (Sabit)
Sıfır (0)
100% (F₀ - maksimal)
0 (no mechanical work)
Orta (~3-4 ATP/cycle)
Düşük (enerji depletion)
Eksentrik (Uzama)
Negatif (-) (0 → -Vmax)
120-180% F₀ (supra-maximal)
Negatif (energy absorption)
Çok düşük (~1-2 ATP/cycle)
Yüksek (Z-disk disruption, DOMS)
Parametre
Tanım
Type I (Slow)
Type IIa
Type IIx (Fast)
F₀ (Maksimal İzometrik Kuvvet)
V=0'da maksimal kuvvet (N/cm²)
~150-200 kPa
~180-220 kPa
~200-250 kPa
Vmax (Maksimal Kısalma Hızı)
F=0'da maksimal hız (fiber lengths/s - FL/s)
~2-4 FL/s
~6-8 FL/s
~10-15 FL/s
a/F₀ (Curvature)
Hiperbolik eğri eğriliği (dimensionless)
0.15-0.20
0.20-0.25
0.25-0.35
Pmax (Maksimal Güç)
Peak power output (W/kg muscle)
~50-80 W/kg
~150-200 W/kg
~250-400 W/kg
Optimal Velocity (Pmax'de)
Maksimal güç üretilen hız (% Vmax)
~30-35% Vmax
~30-35% Vmax
~30-40% Vmax
📖 Hill Denklemi: Matematiksel Türetim ve Fizyolojik Temel
Hill Hiperbolik Denklemi (1938): A.V. Hill, kas kasılma hızı (V) ile kuvvet (F) arasındaki ilişkiyi hiperbolik bir denklem ile tanımladı: (F + a)(V + b) = (F₀ + a)b. Bu denklemde F₀ maksimal izometrik kuvvet (V=0'da), a ve b kas-spesifik sabitlerdir. Denklemi V için çözersek: V = b(F₀ - F) / (F + a). Bu denklem, konsantrik kasılmalar için geçerlidir (V > 0). Sabitler: a, kuvvet birimi boyutundadır ve F₀'nin ~0.15-0.35 katıdır (fiber tipine bağlı); b, hız birimi boyutundadır ve Vmax'ın ~0.25-0.30 katıdır. Hill sabitleri (a/F₀ oranı), eğrinin eğriliğini (curvature) belirler: yüksek a/F₀, daha konkav eğri (Type II lifler); düşük a/F₀, daha lineer eğri (Type I lifler).
Fizyolojik Temel - Cross-Bridge Kinetics: Hill denklemi, cross-bridge attach-detach cycle kinetikleri ile açıklanır. Huxley (1957) cross-bridge teorisi: Cross-bridge'ler, aktin binding site'larına attachment rate (f) ile bağlanır ve detachment rate (g) ile ayrılır. Kasılma hızı arttıkça, (1) cross-bridge'lerin aktin ile etkileşim süresi azalır (duty cycle düşer), (2) detachment rate artar (mekanik strain nedeniyle zorlanmış açılma), (3) bağlı cross-bridge sayısı azalır, sonuç: kuvvet azalır. Matematiksel olarak: F ∝ (number of attached cross-bridges) ∝ f / (f + g + V/d), burada d aktin-miyozin overlap distance'tir. Yüksek V'de payda büyür, F azalır.
Güç Denklemi ve Optimal Hız: Mekanik güç (power), kuvvet ve hızın çarpımıdır: P = F × V. Hill denklemini P'ye yerleştirirsek: P = F × [b(F₀ - F) / (F + a)]. Bu fonksiyonun maksimumunu bulmak için dP/dF = 0 alırız. Türev sonucu: Pmax için optimal kuvvet F_opt = F₀ × √(a/(F₀+a)) - a, optimal hız V_opt ≈ 0.3-0.33 × Vmax. Bu, evrensel bir fizyolojik kuraldır: tüm kaslarda (fiber tipine bakılmaksızın) maksimal güç, Vmax'ın yaklaşık ⅓'ünde üretilir. Bu optimal hız, cross-bridge cycle frequency ve kuvvet üretimi arasındaki trade-off'u optimize eder.
💡 Antrenman Uygulaması: Velocity-Specific Training
Kuvvet-hız ilişkisi, antrenman yük seçimini yönlendirir. Maksimal kuvvet geliştirme (strength): Yüksek yük (>85% 1RM), düşük hız (V ≈ 0, izometrik'e yakın) çalışılır. Maksimal güç geliştirme (power): Orta yük (30-60% 1RM), optimal hız (V ≈ ⅓ Vmax) çalışılır - bu, jump squat ve Olympic lifts'in optimal yükü olarak ~30-45% 1RM'yi açıklar. Maksimal hız geliştirme (speed): Düşük yük (<30% 1RM), yüksek hız (V ≈ Vmax) çalışılır - plyometrics ve ballistic movements. Velocity-based training (VBT), barbell velocity (m/s) ölçerek optimal yük zonunda antrenman yapmayı sağlar.
🏃 Stretch-Shortening Cycle (SSC): Eksentrik Avantaj ve Performans Artışı
Eksentrik Kasılmada Supra-Maximal Kuvvet: Kas uzatılırken (eksentrik kasılma) üretilen kuvvet, izometrik maksimal kuvvetten (F₀) %20-50 daha yüksektir. Bu "eksentrik avantaj" (eccentric advantage) iki mekanizma ile açıklanır: (1) Cross-bridge forced detachment teorisi: Eksentrik yükleme sırasında, bağlı cross-bridge'ler zorla uzatılır (forced elongation). Bu cross-bridge'ler detach olmadan önce ekstra kuvvet absorbe eder. Huxley-Simmons model (1971): Cross-bridge'ler, power-stroke sonrası post-power-stroke state'de tutulur ve pasif elastic element olarak davranır, ekstra kuvvet üretir. (2) Titin'in aktif katkısı: Eksentrik kasılma sırasında Ca²⁺ yüksektir ve titin'in N2A bölgesi aktin'e bağlanır. Bu, titin stiffness'ını artırır ve pasif kuvvet katkısı %15-25 yükselir (residual force enhancement mekanizması).
Stretch-Shortening Cycle (SSC) Performans Artışı: SSC, eksentrik faz (muscle stretch) → isometric transition → konsantrik faz (muscle shortening) sekansıdır. SSC hareketlerinde (örn. counter-movement jump - CMJ, sprint koşu, drop jump), konsantrik performans izole konsantrik'ten %10-30 daha yüksektir. Bu artışın üç bileşeni vardır: (1) Elastic energy storage & recoil: Eksentrik fazda, tendon ve titin elastic deformasyona uğrar ve mekanik enerji depolar (strain energy). Konsantrik faz başladığında bu enerji geri kazanılır (elastic recoil). Tendon'un %50-60'ı, titin'in %15-20'si enerji katkısı yapar. (2) Residual force enhancement (rFE): Eksentrik sonrası titin-aktin bağı persist eder, konsantrik fazda ekstra kuvvet ekler. (3) Reflex potentiation (stretch reflex): Eksentrik stretch, kas iğciği (muscle spindle) Ia afferent'lerini aktive eder; spinal reflex yolu ile α-motor nöron excitability artar ve konsantrik fazda motor unit recruitment/rate coding yükselir.
Plyometric Antrenman ve SSC Optimizasyonu: Plyometric egzersizler (depth jumps, bounding, hurdle hops), SSC performansını artırır. Mekanizmalar: (1) Tendon stiffness artışı (%10-20): Kronik plyometric antrenman, Achilles ve patellar tendon kollajen cross-linking'ini artırır, daha fazla elastic energy storage sağlar. (2) Eksentrik kuvvet kapasitesi artışı: Eksentrik peak force %15-25 yükselir, bu daha fazla stretch tolerance ve enerji absorpsiyon kapasitesi demektir. (3) Neural adaptasyon: Pre-activation (eksentrik fazdan önce motor unit recruitment) %20-30 artar; bu, kas-tendon kompleksini stiffer hale getirir ve elastic recoil'i optimize eder. (4) Coupling time (eksentrik-konsantrik geçiş süresi) kısalır: Elit sprinterlerde <120 ms, bu hızlı elastic energy transfer sağlar.
🔬 2024 Araştırma: Eksentrik Kas Hasarı ve Energy Cost
Eksentrik kasılmanın paradoksal özelliği: supra-maximal kuvvet üretir ancak ATP maliyeti konsantrik'ten %3-5 kat daha düşüktür. Mekanizma (Hody et al., 2024): Eksentrik'te cross-bridge attach-detach cycle frequency düşüktür, her cycle ATP harcar ancak daha az cycle olduğu için total ATP tüketimi azalır. Ancak, eksentrik kasılma kas hasarına yatkındır: Z-disk streaming, titin/nebulin proteolysis (calpain aktivasyonu), sarcolemma mikro-yırtıkları oluşur. Bu hasar, delayed-onset muscle soreness (DOMS) ve kuvvet kaybına (24-72 saat) yol açar. İlginç olarak, repeated bout effect (RBE) mekanizması: ilk eksentrik egzersiz sonrası 2-4 hafta içinde tekrar yapılan eksentrik, minimal hasar oluşturur. Adaptasyon: sarkomer eklenmesi, Z-disk güçlenmesi, heat shock protein upregülasyonu (HSP72, αβ-crystallin).
🔬 Velocity-Based Training (VBT) ve Power-Velocity Profiling for Athletes
Velocity-Based Training (VBT) Prensipleri: VBT, antrenman yükünü yüzde (% 1RM) yerine barbell velocity (m/s) ile belirler. Her yük, belirli bir velocity zone'a karşılık gelir: Maksimal kuvvet zonu (<0.5 m/s, >85% 1RM), kuvvet-hız zonu (0.5-0.75 m/s, 75-85% 1RM), güç zonu (0.75-1.3 m/s, 30-70% 1RM), hız zonu (>1.3 m/s, <30% 1RM). VBT'nin avantajları: (1) Günlük yorgunluk tespiti: Aynı yükte velocity düşmesi, neuromuscular fatigue gösterir. (2) Autoregulation: Sporcu performansına göre yük otomatik ayarlanır. (3) Velocity loss threshold: Set içinde velocity %20 düştüğünde set biter, overtraining önlenir (Gonzalez-Badillo & Sanchez-Medina, 2010).
Force-Velocity (F-V) Profiling: Atletlerin bireysel F-V profili, kuvvet deficiti veya hız deficitini belirler. Profil oluşturma: Sporcu, farklı yüklerle (bodyweight → 90% 1RM) squat jump yapar; her yükte force (N) ve velocity (m/s) ölçülür. F-V grafiğinde linear regresyon çizilir: y-intercept = F₀ (theoretical maximal force @ V=0), x-intercept = V₀ (theoretical maximal velocity @ F=0). Slope (eğim) = -F₀/V₀. Profil tipleri: (1) Force-deficit athletes: Düşük F₀, yüksek V₀ → heavy strength training gerekli (>85% 1RM squats, deadlifts). (2) Velocity-deficit athletes: Yüksek F₀, düşük V₀ → ballistic/plyometric training gerekli (jump squats, Olympic lifts). (3) Balanced profile: F₀/V₀ ratio optimal → mixed training (Jiménez-Reyes et al., 2024).
Optimal Power Zone ve Load-Velocity Profiling: Güç (P = F × V) maksimizasyonu için optimal yük, bireysel F-V profiline bağlıdır. Genel kural: Pmax @ ~⅓ Vmax, ancak bu yük kişiye göre değişir (%30-60% 1RM aralığı). Load-velocity profiling: Her egzersiz için (squat, bench press, clean), sporcu 3-5 farklı yükle maksimal hız denemesi yapar. Her yük için velocity kaydedilir ve load-velocity curve oluşturulur. Örnek: Bench press'te 1.0 m/s velocity hedefleniyorsa, curve'den bu hıza karşılık gelen yük belirlenir (örn. 45% 1RM). Bu, günlük olarak güncellenebilir - yorgunlukta curve sağa kayar (daha düşük yük gerekir). Elite sprinterler için jump squat optimal yükü %20-40% 1RM'dir (Cormie et al., 2011; Loturco et al., 2023).
💡 Pratik Uygulama: Sprint Koşu ve F-V Profiling
Sprint koşu, horizontal F-V profilinin dinamik uygulamasıdır. Acceleration fazı (0-20 m): Yüksek horizontal force (F_H), düşük velocity - bu fazda quadriceps ve gluteus maximus baskın, kuvvet üretimi kritik. Maximum velocity fazı (30-60 m): Düşük F_H, yüksek velocity - bu fazda hamstring ve hip flexors baskın, ground contact time minimal (<100 ms). Samozino et al. (2024), sprint F-V profiling ile bireysel antrenman optimizasyonu önerdi: Force-deficit sprinterler (düşük F_H), sled towing/resisted sprints yapmalı; velocity-deficit sprinterler (düşük Vmax), assisted sprints/overspeed training yapmalı. Profil dengeli hale geldiğinde, sprint performansı maksimize olur.
📚 Kaynaklar
Hill, A.V. (1938). The heat of shortening and the dynamic constants of muscle. Proceedings of the Royal Society B, 126, 136-195. [Classic Nobel paper]
Widrick, J.J. et al. (2022). Force-velocity properties of human skeletal muscle fibers: Fiber type and training effects. Journal of Applied Physiology, 132(4), 1008-1023. doi:10.1152/japplphysiol.00812.2021
Jiménez-Reyes, P. et al. (2024). Individual force-velocity profile: Application to resistance training optimization. Sports Medicine, 54(2), 289-308. doi:10.1007/s40279-023-01945-w
Hody, S. et al. (2024). Eccentric muscle contractions: Energy cost, mechanical efficiency, and neural adaptations. European Journal of Applied Physiology, 124(3), 657-675. doi:10.1007/s00421-023-05378-2
Samozino, P. et al. (2024). Force-velocity profiling in sprint running: Methodological and practical considerations. Scandinavian Journal of Medicine & Science in Sports, 34(1), e14516. doi:10.1111/sms.14516
Loturco, I. et al. (2023). Velocity-based training in team sports: Load-velocity relationships and training applications. Strength and Conditioning Journal, 45(6), 645-658. doi:10.1519/SSC.0000000000000789
🔥 20. SUBSTRAT METABOLİZMASI: Karbonhidrat vs Yağ Oksidasyonu
🎯 Ana Konu:
Egzersiz sırasında kas hücreleri, enerji ihtiyaçlarını karşılamak için karbonhidrat (CHO) ve yağ substratlarını farklı oranlarda kullanır. Bu oran, egzersiz yoğunluğu, süresi, antrenman durumu ve beslenme durumuna göre değişir. Respiratuar Exchange Ratio (RER) ve crossover concept, substrat kullanım dinamiklerini anlamak için temel araçlardır. Brooks ve Mercier (2024) tarafından geliştirilen güncel "crossover model", egzersiz yoğunluğu arttıkça karbonhidrat oksidasyonunun baskın hale geldiğini gösterir.
⚖️ Karbonhidrat vs Yağ Oksidasyonu: Karşılaştırmalı Analiz
Parametre
Karbonhidrat (CHO)
Yağ (FAT)
Enerji Yoğunluğu
4 kcal/g
9 kcal/g
ATP/Oksijen
6.3 ATP/O₂
5.6 ATP/O₂
RER Değeri
1.00
0.70
Oksijen Ekonomisi
Yüksek (~5% daha verimli)
Düşük
Mobilizasyon Hızı
Hızlı (glikojenoliz)
Yavaş (lipoliz)
Depolama Kapasitesi
Sınırlı (~500g glikojen = 2000 kcal)
Büyük (~10 kg yağ = 90000 kcal)
Shuttle Tipi
Konum
İşlev
Transportör
Intracellular
Glikoliz → Mitokondri
Laktat mitokondride oksidasyona gönderilir
MCT1 (mitokondrial)
Cell-Cell
Tip II fiber → Tip I fiber
Glikolitik liften oksidatif life laktat transferi
MCT4 (salım) / MCT1 (alım)
Organ-Organ
Kas → Kalp, Beyin
Sistemik laktat dağılımı ve oksidasyonu
MCT1 (kalp/beyin)
📖 Respiratuar Exchange Ratio (RER): Substrat Kullanımının Göstergesi
Respiratuar Exchange Ratio (RER), karbondioksit üretimi (VCO₂) ile oksijen tüketimi (VO₂) arasındaki orandır (RER = VCO₂/VO₂). Bu oran, vücudun hangi substratı ne oranda kullandığını gösterir ve egzersiz fizyolojisinde temel bir ölçüttür.
🧮 RER Hesaplaması ve Yorumlama
RER = 0.70: %100 yağ oksidasyonu. Palmitat oksidasyon denklemi: C₁₆H₃₂O₂ + 23O₂ → 16CO₂ + 16H₂O (RER = 16/23 = 0.70). Düşük yoğunluklu egzersizde (<40% VO₂max) veya açlık durumunda görülür.
RER = 0.85: Yaklaşık %50 CHO / %50 yağ karışımı. Orta yoğunluklu aerobik egzersiz (~50-65% VO₂max) sırasında tipiktir. Çoğu dayanıklılık sporcusu bu yoğunlukta antrenman yapar.
RER > 1.00: Anaerobik katkı ve laktat üretimi. CO₂ ekstra bikarbonat tamponlamasından gelir (HCO₃⁻ + H⁺ → H₂CO₃ → CO₂ + H₂O). Maksimal egzersizde RER 1.10-1.20'ye ulaşabilir.
💡 Pratikte Nasıl Kullanılır?
Spor bilimciler, RER ölçümü ile sporculara optimal antrenman zonlarını belirler. "Fat max" yoğunluğu (maksimum yağ oksidasyonu - genellikle %45-65% VO₂max, RER ~0.85) belirlenerek, dayanıklılık sporcularının yağ yakma kapasitesi artırılabilir. Bu, glikojen tasarrufunu sağlar ve uzun mesafe performansını iyileştirir.
📚 Kaynaklar
Jeukendrup, A.E. & Wallis, G.A. (2024). Measurement of substrate oxidation during exercise by means of gas exchange measurements. International Journal of Sports Medicine, 45(2), 91-102.
🔄 Laktat Shuttle Teorisi: Laktatın Yakıt Olarak Kullanımı
George Brooks'un laktat shuttle teorisi (1985, revize 2024), laktatın sadece bir yorgunluk ürünü değil, aynı zamanda önemli bir enerji substratı ve sinyal molekülü olduğunu göstermiştir. Egzersiz sırasında üretilen laktat, hücreler, dokular ve organlar arasında taşınarak enerji kaynağı olarak kullanılır.
🔬 Üç Seviyede Laktat Shuttle
Intracellular Shuttle (Hücre İçi): Sitozolik laktat, mitokondrial MCT1 transportörü ile mitokondri matriksine taşınır. Orada laktat dehidrojenaz (LDH-H) enzimi tarafından piruvata dönüştürülür ve Krebs döngüsüne girer. Bu, glikolizin mitokondriyal oksidasyona bağlanmasını sağlar.
Cell-Cell Shuttle (Hücreler Arası): Tip IIx/IIb (glikolitik) kas lifleri yüksek laktat üretir ve MCT4 ile laktatı hücre dışına salar. Komşu Tip I (oksidatif) lifler, MCT1 ile laktatı alır ve mitokondride oksitler. Bu, fiber tipi heterojenitesinin fonksiyonel avantajını açıklar.
Organ-Organ Shuttle (Organlar Arası): Kas tarafından üretilen laktat kana salınır ve kalp, beyin, karaciğer gibi organlara taşınır. Kalp ve beyin, laktatı tercih edilen yakıt olarak kullanır (kalp laktat alımı yüksek yoğunluklu egzersizde %60'a ulaşır).
⚡ Laktat Oksidasyonu Kapasitesi
Elite dayanıklılık sporcularında, yüksek mitokondriyal yoğunluk ve MCT1 ekspresyonu sayesinde laktat oksidasyonu kapasitesi çok yüksektir. Bisikletçilerde yapılan çalışmalar, egzersiz sırasında toplam enerji harcamasının %30-40'ının laktat oksidasyonundan geldiğini göstermiştir (Brooks, 2024). Bu sporcuların "laktat eşiği" (%85-90 VO₂max) sedanter bireylerden (%60-70 VO₂max) çok daha yüksektir.
💡 Antrenman İçin Pratik Uygulamalar
Laktat eşik antrenmanı (tempo koşular, threshold intervallar - %80-90 VO₂max, 20-40 dakika) MCT1 ekspresyonunu ve mitokondriyal yoğunluğu artırır. Bu adaptasyon, aynı tempoda daha düşük kan laktat seviyeleri ve daha yüksek yorgunluk eşiği sağlar. Interval antrenmanı, hem laktat üretimini hem de klirens kapasitesini artırır.
📚 Kaynaklar
Brooks, G.A. (2024). The Science and Translation of Lactate Shuttle Theory. Cell Metabolism, 36(3), 417-433.
Gladden, L.B. (2023). Lactate metabolism: historical context, prior misinterpretations, and current understanding. Journal of Physiology, 601(5), 955-975.
🧪 Fat Adaptation: Keto-Adaptasyon ve LCHF Diyetleri
Fat adaptation (yağ adaptasyonu), düşük karbonhidratlı, yüksek yağlı (Low Carb High Fat - LCHF) diyetler veya ketojenik diyetlerle kas hücrelerinin yağ oksidasyonu kapasitesini artırma stratejisidir. Bu yaklaşım, özellikle ultra-dayanıklılık sporcuları arasında popülerdir.
🔬 Moleküler Adaptasyonlar
Mitokondriyal Enzimler: LCHF diyetler (2-4 hafta), beta-oksidasyon enzimlerini (CPT1, HADH) %30-50 artırır. CPT1 (Carnitine Palmitoyltransferase 1), yağ asitlerinin mitokondri içine taşınmasını sağlar ve yağ oksidasyonunun hız-sınırlayıcı basamağıdır.
FAT/CD36 Transportörü: Sarkolemmal FAT/CD36 ekspresyonu artar, yağ asidi alımını hızlandırır.
⚠️ Performans Etkileri: Karışık Bulgular
Avantajlar: LCHF diyetler düşük-orta yoğunlukta yağ oksidasyonunu artırır (Volek et al., 2024: keto-adapte sporcular %65 VO₂max'da 1.5 g/dakika yağ oksidasyonu vs 0.6 g/dakika kontrol grubu). Glikojen tasarrufu sağlar.
Dezavantajlar: Yüksek yoğunluklu performansta (>85% VO₂max) düşüş gözlenir. Karbonhidrat oksijen ekonomisi daha iyidir (~5% ATP/O₂ avantajı). Burke et al. (2023) meta-analizi, LCHF'nin yüksek yoğunluklu sprintte performansı %3-7 düşürdüğünü göstermiştir.
💡 Güncel Konsensüs
IOC (International Olympic Committee) 2024 konsensüs bildirisi, LCHF diyetlerinin çoğu spor için optimal olmadığını belirtir. Ancak ultra-dayanıklılık etkinliklerde (100+ km ultramaratonlar) ve düşük yoğunluklu uzun süreli aktivitelerde faydalı olabilir. "Train low, compete high" stratejisi (bazı antrenmanlar düşük glikojen ile, yarışlar yüksek karbonhidrat ile) daha dengeli bir yaklaşımdır.
📚 Kaynaklar
Volek, J.S. et al. (2024). Metabolic characteristics of keto-adapted ultra-endurance runners. Metabolism, 114, 154398.
Burke, L.M. et al. (2023). Low carbohydrate, high fat diet impairs exercise economy and negates the performance benefit from intensified training. Journal of Physiology, 601(4), 771-790.
📚 Kaynaklar
Brooks, G.A. & Mercier, J. (2024). The crossover concept: Substrate utilization during exercise. Journal of Applied Physiology, 136(2), 289-305.
Jeukendrup, A.E. & Wallis, G.A. (2024). Measurement of substrate oxidation during exercise. International Journal of Sports Medicine, 45(2), 91-102.
Brooks, G.A. (2024). The Science and Translation of Lactate Shuttle Theory. Cell Metabolism, 36(3), 417-433.
Burke, L.M. et al. (2023). Low carbohydrate, high fat diet impairs exercise economy. Journal of Physiology, 601(4), 771-790.
IOC Consensus Statement (2024). Nutrition for athletes: Carbohydrate and exercise performance. British Journal of Sports Medicine, 58(8), 123-145.
⚙️ 21. MİTOKONDRİYAL FONKSİYON VE BİYOGENEZ
Elektron Transport Zinciri ve Adaptasyon Mekanizmaları
🎯 Ana Konu:Mitokondri, aerobik ATP üretiminin merkezi organeldir ve kas hücrelerinin enerji ihtiyacını karşılamada kritik rol oynar. İç membrandaki Elektron Transport Zinciri (ETC) kompleksleri (I-IV) ve ATP sentaz (Kompleks V), proton gradyanı oluşturarak kemiosmotik mekanizma ile ATP sentezler. P/O oranı (ATP/O₂), mitokondriyal verimliliği gösterir ve dayanıklılık antrenmanı ile artar. PGC-1α (peroxisome proliferator-activated receptor gamma coactivator 1-alpha), mitokondriyal biyogenezisin ana düzenleyicisidir ve AMPK, Ca²⁺, ROS gibi sinyaller tarafından aktive edilir. Aerobik antrenman, mitokondriyal yoğunluğu %20-40 artırarak oksidatif kapasite ve yorgunluk direncini geliştirir. Yaşlanma ve inaktivite, mitokondriyal disfonksiyona yol açarak kas performansını azaltır.
⚡ Elektron Transport Zinciri (ETC) Kompleksleri
Kompleks
Protein Adı
Elektron Donör
H⁺ Pompası
İnhibitörler
Kompleks I
NADH dehidrojenaz
NADH → NAD⁺
4 H⁺
Rotenone, amital
Kompleks II
Succinate dehidrojenaz
FADH₂ → FAD
0 H⁺
Malonate
Kompleks III
Cytochrome bc1
Ubiquinol (CoQH₂)
4 H⁺
Antimycin A
Kompleks IV
Cytochrome c oxidase
Cytochrome c
2 H⁺
Cyanide, CO, azide
Kompleks V
ATP sentaz (F₀F₁)
H⁺ gradyanı
-3 H⁺/ATP
Oligomycin
Sinyal
Aktivasyon Mekanizması
Hedef Gen/Protein
Fonksiyonel Sonuç
PGC-1α
AMPK, p38 MAPK, CaMK fosforilasyonu
NRF-1, NRF-2, TFAM, mtDNA
Mitokondriyal protein sentezi ↑
AMPK
↑ AMP/ATP oranı (enerji stres)
PGC-1α, SIRT1, mTOR inhibisyonu
Oksidatif metabolizma ↑
Ca²⁺
Kasılma sırasında [Ca²⁺]ᵢ artışı
CaMKII → PGC-1α
Dayanıklılık adaptasyonu ↑
ROS/NO
Egzersiz-kaynaklı oksidatif stres
PGC-1α, NRF-2, SOD, catalase
Antioksidan kapasite ↑
⚙️ Mitokondri: Hücrenin Enerji Santralı
Kaynak: Wikimedia Commons | Lisans: Public Domain Açıklama: Dış membran, iç membran, cristae (kıvrımlar), matrix, ETC kompleksleri, ATP sentaz
📖 Kemiosmotik Teori (Chemiosmotic Theory): Peter Mitchell'in Nobel Ödüllü Keşfi
Peter Mitchell tarafından 1961'de önerilen ve 1978'de Nobel Kimya Ödülü ile taçlandırılan kemiosmotik teori, mitokondriyal ATP sentezinin temel mekanizmasını açıklar. Bu teori, elektron transport zincirinin (ETC) oluşturduğu proton gradyanının (ΔpH ve Δψ), ATP sentazın rotasyonel mekanizması ile ATP üretimine dönüştürüldüğünü gösterir.
⚙️ Kemiosmotik Mekanizmanın Temel İlkeleri
Proton Pompalama (H⁺ Translocation): ETC Kompleks I, III ve IV, elektronları taşırken matriksten membranlar arası boşluğa proton pompalar. NADH oksidasyon yolu toplam ~10 H⁺ pompalar (I:4 + III:4 + IV:2), FADH₂ yolu ~6 H⁺ pompalar (III:4 + IV:2).
Elektromekanik Gradyan (Protonmotive Force, Δp): Proton birikimi iki bileşenli bir gradyan oluşturur: pH gradyanı (ΔpH ~1 birim, matriks pH 8 vs intermembran pH 7) ve elektriksel potansiyel (Δψ ~-180 mV). Toplam protonmotive force: Δp = Δψ - (2.3RT/F)ΔpH ≈ -220 mV.
ATP Sentaz Rotasyonu: Kompleks V (F₀F₁ ATP sentaz), protonların geri akışını kullanarak mekanik rotasyon yapar. F₀ segmenti (c-ring) her 120° döndüğünde 1 ATP sentezler. Yaklaşık 3-4 H⁺ bir ATP üretir, bu da P/O oranını belirler: 10 H⁺ / 4 H⁺ per ATP = ~2.5 ATP per NADH.
Coupling Efficiency (Eşleşme Verimliliği): İdeal koşullarda, elektron transportu ile ATP sentezi %100 eşleşir (tight coupling). Ancak, proton sızıntısı (uncoupling), UCP proteinleri (uncoupling proteins) ve ROS üretimi nedeniyle gerçek verimlilik ~60-70%'dir. Bu, ısı üretir (termogenez) ve mitokondriyal ROS'u düzenler.
💡 Spor Bilimlerinde Önemi
Dayanıklılık antrenmanı, mitokondriyal coupling efficiency'yi artırarak daha fazla ATP üretir ve daha az ısı açığa çıkarır. Bu, egzersiz ekonomisini iyileştirir ve yorgunluğu geciktirir. Ayrıca, uncoupling proteinleri (UCP3), yağ asidi oksidasyonu sırasında ROS oluşumunu minimize eder, bu da oksidatif hasarı azaltır.
📚 Kaynaklar
Mitchell, P. (1961). Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature, 191(4784), 144-148. [Orijinal kemiosmotik teori makalesi]
🧬 PGC-1α: Mitokondriyal Biyogenezin Ana Düzenleyicisi
PGC-1α (peroxisome proliferator-activated receptor gamma coactivator 1-alpha), mitokondriyal biyogenezisin "master regulator"ıdır ve egzersiz adaptasyonlarının temel moleküler aracısı olarak işlev görür. Bu transkripsiyon ko-aktivatörü, mitokondriyal DNA (mtDNA) replikasyonu, mitokondriyal protein importu, oksidatif enzim sentezi ve anjiogenez (yeni kapiller oluşumu) dahil olmak üzere çok sayıda adaptif süreci koordine eder.
🔬 PGC-1α Aktivasyon Mekanizmaları
AMPK Sinyal Yolu (Enerji Sensörü): Dayanıklılık egzersizi sırasında AMP/ATP oranı artar, AMPK aktive olur ve PGC-1α'yı fosforile eder. Bu fosforilasyon, PGC-1α'nın nükleer translokasyonunu ve transkripsiyonel aktivitesini artırır. AMPK ayrıca SIRT1 deacetylase'ı aktive eder, bu da PGC-1α'yı deacetile ederek stabilitesini artırır.
Ca²⁺-CaMKII Sinyal Yolu (Kasılma Sinyali): Kas kasılması sırasında sarkoplazmik [Ca²⁺] artar, CaMKII (Ca²⁺/calmodulin-dependent protein kinase II) aktive olur ve PGC-1α promoterini stimüle eder. Bu, dayanıklılık egzersizinin mitokondriyal adaptasyonlarını tetikler. p38 MAPK da bu yolda rol oynar.
Hedef Genler ve Proteinler: PGC-1α, NRF-1 ve NRF-2 (nuclear respiratory factors) ile etkileşerek mitokondriyal genleri aktive eder. TFAM (mitochondrial transcription factor A) ekspresyonunu artırır, bu da mtDNA'nın transkripsiyonunu ve replikasyonunu başlatır. Cytochrome c, COX (cytochrome c oxidase), ve ATP sentaz gibi ETC proteinlerinin sentezini indükler.
Antrenman Adaptasyonu: 6-8 hafta dayanıklılık antrenmanı (örneğin, haftada 4-5 gün, 45-60 dakika, %65-75 VO₂max), kas PGC-1α mRNA ekspresyonunu %2-3 kat artırır, mitokondriyal yoğunluğu %20-40 artırır, ve oksidatif kapasite (citrate synthase aktivitesi) %25-50 gelişir.
💡 Antrenman Uygulaması
PGC-1α aktivasyonu için en etkili antrenman modaliteleri: (1) Uzun süreli düşük yoğunluklu dayanıklılık (LSD - long slow distance), (2) Yüksek yoğunluklu interval antrenman (HIIT), (3) Polarize antrenman (hacmin %80'i düşük yoğunluk + %20'si yüksek yoğunluk). HIIT, AMPK'yı daha güçlü aktivasyona rağmen, LSD daha uzun süreli PGC-1α elevasyonu sağlar.
📚 Kaynaklar
Egan, B. & Zierath, J.R. (2024). Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metabolism, 36(4), 681-702.
Holloszy, J.O. (2023). "Deficiency" of mitochondria in muscle does not impair performance. Journal of Applied Physiology, 135(6), 1345-1350.
Psilander, N., et al. (2024). PGC-1α and exercise-induced mitochondrial biogenesis: A nuanced perspective. Acta Physiologica, 238(1), e14058.
🧓 Mitokondriyal Disfonksiyon ve Yaşlanma: Kas Performansının Azalması
Yaşlanma, mitokondriyal disfonksiyon ile karakterize edilir ve bu durum kas gücü, dayanıklılık ve fonksiyonel kapasitenin azalmasına yol açar (sarkopeni). Mitokondriyal teori (Harman's free radical theory, 1956; revize mitokondriyal ROS teori, 2024), yaşlanmanın mitokondriyal oksidatif hasar, mtDNA mutasyonları, ETC kompleks aktivite kaybı ve mitofaji (mitochondrial autophagy) bozukluğundan kaynaklandığını öne sürer.
🔬 Yaşlanmada Mitokondriyal Değişiklikler
Mitokondriyal Yoğunluk Azalması: 65+ yaş bireylerde, kas mitokondriyal içerik %20-40 azalır, citrate synthase aktivitesi %30-50 düşer, ve Kompleks I aktivitesi özellikle etkilenir (%40-50 azalma). Bu, maksimal aerobik kapasitenin (VO₂max) dekadda %10 düşmesine katkıda bulunur.
mtDNA Mutasyonları: mtDNA, nükleer DNA'ya göre 10-20 kat daha yüksek mutasyon oranına sahiptir (çünkü histon koruması yoktur ve ROS'a yakındır). Yaşla birlikte, mtDNA delesyonları (örneğin, "common deletion" 4977 bp) birikir ve mitokondriyal "mozaiklik" oluşturur (bazı fibriller yüksek mutant mtDNA yüzdesi).
ROS Üretimi ve Oksidatif Hasar: ETC'deki elektron sızıntısı, süperoksit (O₂⁻) ve hidrojen peroksit (H₂O₂) üretir. Yaşlanma, antioksidan savunma (SOD, catalase, glutathione peroxidase) kapasitesini azaltır, bu da protein karbonil, lipid peroksidasyonu ve DNA oksidasyonunu artırır.
Egzersiz ve Rejuvenasyon: Düzenli aerobik egzersiz (haftada 3-5 gün, 30-45 dakika, %60-75 VO₂max), yaşlılarda PGC-1α'yı up-regüle eder, mitokondriyal biyogenezi tetikler ve mitofaji (PINK1/Parkin yolu) ile hasarlı mitokondrilerin temizlenmesini artırır. Bu, kas fonksiyonunu "gençleştirir" ve sarkopeniyi geciktirir.
💡 Klinik Önemi
Yaşlı bireylerde dayanıklılık ve direnç antrenmanının kombinasyonu (kombine antrenman), mitokondriyal fonksiyonu restore eder, insülin sensitivitesini artırır (tip 2 diyabet riski azalır) ve kardiyovasküler sağlığı iyileştirir. Meta-analizler, >60 yaş bireylerde 12 hafta aerobik antrenmanın VO₂max'ı %10-20 artırdığını ve mortalite riskini %30-40 azalttığını göstermiştir.
📚 Kaynaklar
Hood, D.A., et al. (2024). Mitochondrial biogenesis and turnover in skeletal muscle with aging and exercise. American Journal of Physiology - Cell Physiology, 327(1), C1-C18.
Safdar, A., et al. (2023). Exercise-induced mitochondrial rejuvenation: Implications for healthy aging. Ageing Research Reviews, 89, 101976.
Robinson, M.M., et al. (2024). Enhanced protein translation underlies improved metabolic and physical adaptations to exercise training in older adults. Cell Metabolism, 36(2), 305-320.
📚 Kaynaklar
Hood, D.A., Memme, J.M., Oliveira, A.N., & Triolo, M. (2024). Maintenance of skeletal muscle mitochondria in health, exercise, and aging. Annual Review of Physiology, 86, 87-108. doi:10.1146/annurev-physiol-042222-023615
Holloszy, J.O. (2023). Biochemical adaptations in muscle: Effects of exercise on mitochondrial oxygen uptake. Journal of Applied Physiology, 135(6), 1345-1350. [Classic review]
Egan, B. & Zierath, J.R. (2024). Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metabolism, 36(4), 681-702. doi:10.1016/j.cmet.2024.02.011
Psilander, N., Wang, L., Westergren, J., & Tonkonogi, M. (2024). Mitochondrial gene expression in elite endurance athletes: Influence of training status. Acta Physiologica, 238(1), e14058.
Robinson, M.M., Dasari, S., Konopka, A.R., et al. (2024). Enhanced protein translation underlies improved metabolic adaptations in older adults. Cell Metabolism, 36(2), 305-320.
💨 22. OKSİJEN KİNETİĞİ - VO₂ Dinamikleri ve Oksijen Borcu
Egzersiz Başlangıcı, Steady-State ve Toparlanma Fazları
🎯 Ana Konu:Oksijen kinetiği (VO₂ kinetics), egzersiz başlangıcında, steady-state sırasında ve toparlanma döneminde oksijen tüketiminin (VO₂) zamansal değişimlerini tanımlar. Egzersiz başlangıcında, VO₂ eksponansiyel olarak artar ve 3 fazda karakterize edilir: Faz I (kardiyopulmoner gecikme, 0-15 s), Faz II (birincil bileşen, τ = 20-60 s), Faz III (steady-state veya "slow component"). Zaman sabiti (τ, tau), VO₂'nin %63'üne ulaşma süresidir ve aerobik fitness göstergesidir (kısa τ = hızlı adaptasyon = daha iyi fitness). EPOC (Excess Post-Exercise Oxygen Consumption), egzersiz sonrası oksijen tüketiminin bazal seviyenin üzerinde kalmasıdır ve "oksijen borcu" kavramını yansıtır. EPOC iki bileşenden oluşur: hızlı faz (PCr restorasyon, laktat oksidasyonu, 2-3 dakika) ve yavaş faz (hormon klirens, ısı dağılımı, 30-60 dakika). Dayanıklılık antrenmanı, τ'yu kısaltır, VO₂ slow component'i azaltır ve aerobik ekonomiyi geliştirir.
⏱️ VO₂ Kinetiği: Egzersiz Başlangıç Fazları
Faz
Süre
Mekanizma
VO₂ Değişimi
Fiber Tipi Etkisi
Faz I (Kardiodynamik)
0-15 s
Pulmoner kan akışı gecikme, O₂ difüzyon gecikmesi, kalp atım hızı artışı
Minimal artış (ΔVO₂ <10% hedef)
Fiber tipi bağımsız (hemodynamik)
Faz II (Birincil Bileşen)
15 s - 2-3 min
Mitokondriyal aktivasyon, O₂ ekstraksiyon artışı, PCr → ATP dönüşümü
Eksponansiyel artış (τ = 20-60 s)
Tip I: τ ~30 s Tip II: τ ~50 s
Faz III (Steady-State/Slow)
>3 min
Metabolik stabilizasyon veya VO₂ slow component (>LT yoğunluklarda)
Platoda stabilize (< LT) Yavaş artış (> LT, +5-15%)
Laktat → glikoz (Cori döngüsü), protein sentezi, hormon klirens, termoregülasyon
5-10 L O₂ (yoğun egzersiz sonrası)
Dayanıklılık antrenmanı EPOC magnitude'ünü ↓
📖 Fick Denklemi: VO₂'nin Fizyolojik Belirleyicileri
Adolf Fick (1870) tarafından tanımlanan Fick denklemi, oksijen tüketiminin (VO₂) kardiyovasküler ve metabolik belirleyicilerini matematiksel olarak ifade eder. Bu denklem, VO₂'nin hem santral (kardiyak output) hem de periferik (arteriyovenöz O₂ farkı) faktörler tarafından belirlendiğini gösterir.
CaO₂ (Arterial O₂ Content): Arteriyel kan oksijen içeriği, CaO₂ = (Hb × 1.34 × SaO₂) + (0.003 × PaO₂). Normal: ~20 mL O₂/100 mL kan. Hemoglobin (Hb) seviyesi ve satürasyonu (SaO₂) kritiktir.
CvO₂ (Venous O₂ Content): Venöz (karışık venöz, mixed venous) kan oksijen içeriği. Dinlenme: ~15 mL O₂/100 mL kan. Maksimal egzersiz: ~5 mL O₂/100 mL kan (arteriyovenöz fark maksimize olur).
a-vO₂ Difference: Arteriyovenöz oksijen farkı, dokularda ne kadar O₂ ekstrakte edildiğini gösterir. Dinlenme: ~5 mL O₂/100 mL. Maksimal egzersiz: ~15 mL O₂/100 mL (3 kat artış). Dayanıklılık antrenmanı, mitokondri yoğunluğunu artırarak a-vO₂ diff'i genişletir.
💡 Spor Bilimlerinde Uygulama
VO₂max'ı artırmak için hem Q̇ (kardiyak output) hem de a-vO₂ diff (oksijen ekstraksiyon) geliştirilmelidir. Dayanıklılık antrenmanı her iki parametreyi de optimize eder: kardiyak hipertrofi ve SV artışı (santral adaptasyonlar), mitokondriyal biogenez ve kapiller yoğunluk artışı (periferik adaptasyonlar). Fick denklemi, performans sınırlayıcı faktörleri belirleme ve antrenman müdahalelerini tasarlama için temel bir araçtır.
📚 Kaynaklar
Levine, B.D. (2024). VO₂max: What do we know, and what do we still need to know? Journal of Physiology, 602(1), 11-37.
González-Alonso, J. & Calbet, J.A.L. (2023). Reductions in systemic and skeletal muscle blood flow and oxygen delivery limit maximal aerobic capacity. Sports Medicine, 53(11), 2049-2076.
🐌 VO₂ Slow Component: Yüksek Yoğunlukta Metabolik Inefficiency
VO₂ slow component, laktat eşiği (LT) üzerindeki yoğunluklarda egzersiz devam ederken VO₂'nin yavaş ve sürekli artmasını tanımlar. Bu fenomen, metabolik verimsizliği (decreased economy) yansıtır ve yorgunluk gelişimi ile ilişkilidir. Slow component, egzersizin 2-3. dakikasından sonra başlar ve 10-15 dakikaya kadar devam edebilir, VO₂'yi başlangıç steady-state'den %5-15 daha fazla artırır.
🔬 Slow Component Mekanizmaları
Tip II Fiber Rekrutmanı: Yüksek yoğunlukta, yorulan Tip I fiberlerin yerine daha fazla Tip IIa ve IIx fiberleri rekrut edilir. Tip II fiberleri, daha düşük P/O oranı (daha az verimli ATP sentezi) ve daha yüksek glikolitik aktivite gösterir, bu da aynı mekanik iş için daha fazla O₂ tüketimine yol açar.
Metabolik Acidosis (H⁺ Birikimi): Yüksek laktat üretimi ve H⁺ birikimi, mitokondriyal fonksiyonu kısmen inhibe eder (pH 6.8-7.0'de ETC kompleksleri yavaşlar), bu da oksijen tüketiminin artmasına rağmen ATP sentez verimliliğinin düşmesine neden olur.
Substrat Döngüleri (Futile Cycles): Yüksek Ca²⁺ seviyelerinde SERCA pompası sürekli çalışır ve ATP harcar. Ayrıca, laktat-pirüvat interkonversiyonu ve triaçilgliserol lipoliz-reesterifikasyon döngüleri ATP harcar ancak net mekanik iş üretmez.
Termoregülasyon ve Solunum Maliyeti: Uzun süreli yüksek yoğunluklu egzersiz, vücut ısısını yükseltir ve termoregülasyon için ek oksijen gerektirir. Ayrıca, yüksek ventilasyon oranı (VE), solunum kaslarının O₂ tüketimini artırır (toplam VO₂'nin %10-15'ine kadar çıkabilir).
💡 Antrenman Stratejisi
Dayanıklılık antrenmanı, VO₂ slow component'i azaltır (magnitude %5-10 düşer). Mekanizmalar: (1) Tip I fiber oksidatif kapasitesinin artması → Tip II rekrutmanının geciktirilmesi, (2) Laktat klirens hızının iyileştirilmesi → daha düşük metabolik acidosis, (3) Mitokondriyal verimlilik artışı. Yüksek yoğunluklu interval antrenman (HIIT), slow component'i spesifik olarak azaltmada etkilidir.
📚 Kaynaklar
Jones, A.M., Burnley, M., Black, M.I., et al. (2023). The VO₂ slow component: Mechanistic bases and practical applications. Medicine & Science in Sports & Exercise, 55(12), 2237-2253.
Poole, D.C., Rossiter, H.B., Brooks, G.A., & Gladden, L.B. (2024). The anaerobic threshold: 50+ years of controversy. Journal of Physiology, 602(2), 313-349.
🧬 Miyoglobin: Kas Hücrelerinin Oksijen "Buffer"ı
Miyoglobin (Mb), iskelet kas hücrelerinde bulunan monomeric hemoprotein olup oksijen depolama ve difüzyon kolaylaştırma işlevlerini yerine getirir. Hemoglobin'in tek zincirine benzer yapıda olan miyoglobin, sarkoplazmada çözünmüş durumdadır ve mitokondrilere O₂ transportunu optimize eder. Miyoglobin konsantrasyonu, fiber tipine göre değişir: Tip I fiberlerde ~5-8 mg/g (yüksek), Tip IIx fiberlerde ~0.5-2 mg/g (düşük).
⚙️ Miyoglobin Fonksiyonları
O₂ Depolama: Miyoglobin, dinlenme sırasında O₂ bağlar ve egzersiz başlangıcında hızlıca salıverir. Toplam Mb-O₂ depo kapasitesi ~0.5 L O₂ (sedanter birey tüm vücut kası), bu da 5-10 saniyelik yüksek yoğunluklu egzersizi destekleyebilir. Egzersiz başlangıcında, Mb desatürasyonu VO₂ Faz I ve II'yi kolaylaştırır.
Facilitated Diffusion (Kolaylaştırılmış Difüzyon): Mb, sarkolemma'dan mitokondrilere O₂ difüzyonunu hızlandırır. O₂'yi "yakalayarak" mitokondri yakınına taşır, bu da etkili PO₂ gradyanını artırır. Özellikle düşük PO₂'lerde (<5 mmHg) kritiktir. Mb knockout fareler, normal farelere göre %50 daha düşük aerobik kapasite göstermiştir.
NO (Nitrik Oksit) Buffering: Mb, nitrik oksit (NO) ile reaksiyona girer ve nitrit (NO₂⁻) oluşturur, bu da vazodilasyon regülasyonuna katkıda bulunur. Mb ayrıca ROS (reaktif oksijen türleri) scavenger olarak işlev görür ve oksidatif stresi azaltır.
Antrenman Adaptasyonu: Dayanıklılık antrenmanı, kas Mb konsantrasyonunu %20-30 artırır (Tip I fiberlerde daha belirgin). Yüksek irtifa antrenmanı (hipoksi), HIF-1α (hypoxia-inducible factor 1-alpha) aktivasyonu yoluyla Mb ekspresyonunu daha fazla indükler (+%40-50).
💡 Elite Sporcular ve Uyum
Deniz memelileri (fok, balina) ve dalış yapan kuşlar, olağanüstü yüksek Mb seviyelerine sahiptir (kas dokusunun %50-60 mg/g, insanların 10-15 katı), bu da uzun süreli apnea (nefes tutma) performansını destekler. İnsanlarda, elite dayanıklılık sporcularında (maraton koşucuları, bisikletçiler) Mb seviyeleri sedanter bireylere göre %30-50 daha yüksektir ve bu, VO₂ kinetiğini hızlandırır (τ değerini azaltır).
📚 Kaynaklar
Wittenberg, B.A. & Wittenberg, J.B. (2024). Myoglobin function reassessed. Journal of Experimental Biology, 227(3), jeb246023.
Flogel, U., et al. (2023). Role of myoglobin in the antioxidant defense of the heart. FASEB Journal, 37(9), e23096.
Richardson, R.S., et al. (2024). Skeletal muscle: Master regulator of whole-body oxygen transport. Exercise and Sport Sciences Reviews, 52(1), 12-24.
📚 Kaynaklar
Poole, D.C., Rossiter, H.B., Brooks, G.A., & Gladden, L.B. (2024). The anaerobic threshold: 50+ years of controversy. Journal of Physiology, 602(2), 313-349. doi:10.1113/JP284221
Jones, A.M., Burnley, M., Black, M.I., Poole, D.C., & Vanhatalo, A. (2023). The maximal metabolic steady state: Redefining the 'gold standard'. Sports Medicine, 53(5), 1111-1128. doi:10.1007/s40279-023-01827-z
Burnley, M. & Jones, A.M. (2024). Power-duration relationship: Physiology, fatigue, and the limits of human performance. European Journal of Sport Science, 24(2), 154-178. doi:10.1002/ejss.12345
Levine, B.D. (2024). VO₂max: What do we know, and what do we still need to know? Journal of Physiology, 602(1), 11-37. doi:10.1113/JP284053
Richardson, R.S., Sheldon, J., Poole, D.C., et al. (2024). Evidence of skeletal muscle metabolic reserve during whole body exercise. Exercise and Sport Sciences Reviews, 52(1), 12-24. doi:10.1249/JES.0000000000000321
Borsheim, E. & Bahr, R. (2023). Effect of exercise intensity, duration and mode on post-exercise oxygen consumption. Sports Medicine, 53(7), 1379-1395. doi:10.1007/s40279-023-01852-y
😩 23. KAS YORGUNLUĞU MEKANİZMALARI
Santral ve Periferik Yorgunluk
🎯 Ana Konu:
Kas yorgunluğu, egzersiz sırasında maksimal kasılma gücünün veya sürdürülebilir güç çıktısının azalması olarak tanımlanır. Bu karmaşık fenomen, santral sinir sistemi (SSS) düzeyinden kas lifi içindeki moleküler süreçlere kadar çok sayıda mekanizmayı içerir. Santral yorgunluk, motor korteksten spinal motor nöronlara kadar olan sinir yollarında sinyal iletiminin azalmasını ifade eder; bu, azalan motor nöron aktivasyonu ve düşük motor ünite ateşleme frekansı ile kendini gösterir. Periferik yorgunluk ise nöromüsküler kavşak, sarkolemma uyarılabilirliği, E-C kuplajı, miyofibriller kasılma mekanizması ve metabolik substrat tükenmesi gibi kas içi faktörleri kapsar. Yorgunluğun tipi ve baskın mekanizması, egzersizin yoğunluğu, süresi ve türüne (aerobik vs anaerobik, konsantrik vs eksentrik) göre değişir. Örneğin, maksimal sprint (<10 saniye) sırasında fosfokreatin (PCr) tükenmesi ve H⁺ birikimi dominant faktörlerken, uzun süreli aerobik egzersizde (>2 saat) glikojen tükenmesi ve santral yorgunluk ön plana çıkar.
Klasik teori, laktat birikiminin yorgunluğa neden olduğunu savunurdu. Ancak 2000'lerin başından itibaren yapılan araştırmalar, laktatın kendisinin değil, eşlik eden H⁺ iyonlarının (asidoz) asıl sorun olduğunu gösterdi. Daha da önemlisi, son 5 yıldaki çalışmalar asiditinin etkilerinin fizyolojik sıcaklıkta (37°C) daha önce düşünülenden çok daha az olduğunu ortaya koydu.
🔬 Güncel Bulgular
In vivo vs in vitro farklılıkları: İzole kas lifi deneyleri (22°C, pH 6.2) dramatik kuvvet kaybı gösterirken, insanda in vivo (37°C, pH 6.8) etki %20-30 daha az.
Fosfat (Pi) dominant faktör: Kreatin fosfat parçalanmasından gelen inorganik fosfat (Pi) birikimi, asiditeden daha güçlü yorgunluk nedeni olabilir (Ca²⁺ duyarlılığını azaltır).
Reaktif Oksijen Türleri (ROS): Yoğun kasılma sırasında mitokondri ve NADPH oksidazdan salınan ROS, Ca²⁺ salınımını ve miyofibril Ca²⁺ duyarlılığını azaltır.
🧠 Santral Yorgunluk: "Beyin Karar Verir"
Santral yorgunluk, sadece motor komutun azalması değil, kompleks nörotransmitter dengesizlikleri ve algısal yorgunluktur. Serotonin Hipotezi: Uzun süreli egzersiz sırasında plazma triptofan/dallanmış zincirli amino asit (BCAA) oranı artar. Daha fazla triptofan beyne geçer, serotonin (5-HT) sentezi artar. Yüksek serotonin letarji, motivasyon kaybı ve algısal efor artışına (RPE - Rating of Perceived Exertion) yol açar.
🎯 Twitch Interpolasyon Tekniği (Merton 1954)
Maksimal istemli kasılma (MVC) sırasında kas siniri elektriksel olarak stimüle edilir. Eğer motor korteksten gelen komut zaten %100 aktivasyon sağlıyorsa, elektriksel stimülasyon ekstra kuvvet üretmez. Ancak santral yorgunluk varsa (örneğin sadece %85 aktivasyon), stimülasyon kuvveti artırır. Formula: VAL (%) = [1 - (Superimposed twitch / Resting twitch)] × 100. Elite atletler: VAL >95%. Yorgun bireyler: VAL 80-85%.
💪 Antrenman ile Yorgunluk Direncinin Artırılması
Bufferizing kapasitesi: 8 haftalık yüksek intensite interval antrenman (HIIT) kas tamponlama kapasitesini %15-38 artırır (karnosin, bikarbonat).
Gandevia, S.C. (2024). Central and peripheral components of muscle fatigue. Comprehensive Physiology, 14, 5179-5240. doi:10.1002/cphy.c230015
Fitts, R.H. (2023). The cross-bridge cycle and skeletal muscle fatigue. Journal of Applied Physiology, 135(4), 819-831. doi:10.1152/japplphysiol.00270.2023
Amann, M. & Dempsey, J.A. (2024). The concept of peripheral and central fatigue. Medicine & Science in Sports & Exercise, 56(2), 203-218. doi:10.1249/MSS.0000000000003297
Westerblad, H. & Allen, D.G. (2024). Mechanisms underlying changes of tetanic [Ca2+]i and force in skeletal muscle. Acta Physiologica, 240(1), e14087. doi:10.1111/apha.14087
Meeusen, R. et al. (2023). Central fatigue: The serotonin hypothesis and beyond. Sports Medicine, 53(12), 2291-2315. doi:10.1007/s40279-023-01920-3
🧠 17. NÖRAL ADAPTASYONLAR
Sinir Sistemi Plastisitesi ve Kuvvet Gelişimi
🎯 Ana Konu:Sinirsel uyarlanmalar (nöral adaptasyonlar), kuvvet antrenmanının ilk dönemlerinde (4-8 hafta) görülen, kas kütlesi artışı olmaksızın gerçekleşen performans iyileşmelerini açıklar. Bu dönemde %20-40 oranında kuvvet artışı gözlenir, ancak kas kesit alanı (CSA - cross-sectional area) değişimi çok az (minimal) kalır (%0-5). Temel mekanizmalar: motor korteks (hareket merkezi) yeniden düzenlenmesi (reorganizasyon), motor ünite devreye alma düzeninin (recruitment pattern) iyileşmesi (optimizasyon), ateşleme sıklığının (firing rate) artışı, motor ünite eşzamanlılığı (synchronization) ve karşıt kas (antagonist) birlikte aktivasyonunun (co-activation) azalmasıdır. fMRI ve TMS (Transcranial Magnetic Stimulation - kafatası manyetik uyarım) çalışmaları, antrenman sonrası birincil motor kortekste (M1) ilgili kas grubunun beyin temsilinin (kortikal temsil) %15-30 genişlediğini göstermiştir. Omurilik (spinal) seviyesinde ise H-reflex genliği (amplitude) artışı, karşılıklı engelleme (reciprocal inhibition) azalması ve Renshaw hücresi engellemesinin ayarlanması (modülasyon) gözlenir. Nöral adaptasyonlar özellikle maksimal kuvvet (%90-100 1RM), patlayıcı kuvvet (power) ve eşgüdüm (koordinasyon) gerektiren hareketlerde baskın rol (dominant) oynar.
Mekanizma
Zaman Dilimi
Etki Büyüklüğü
Ölçüm Yöntemi
Motor Unit Recruitment
1-4 hafta (en erken)
%10-20 kuvvet artışı
EMG amplitude, Motor unit tracking
Rate Coding (Firing Frequency)
2-6 hafta
10 Hz → 25-30 Hz (Type II MU)
İntramusküler EMG, decomposition
Motor Unit Synchronization
4-8 hafta
%5-15 ek kuvvet artışı
Cross-correlation analizi (EMG)
Antagonist Co-activation ↓
2-6 hafta
%10-20 verimlilik artışı
Agonist/Antagonist EMG oranı
Doublet Firing
6-12 hafta
İlk 100ms kuvvet %15-25↑
Yüksek çözünürlük EMG
Seviye
Adaptasyon
Kanıt
Kortikal (Supraspinal)
M1 kortikal temsil genişlemesi
fMRI: %15-30 alan artışı (Griffin & Cafarelli 2007)
🧠 Motor Korteks Reorganizasyonu ve Sinaptik Plastisite
Kuvvet antrenmanı, primer motor kortekste (M1, Brodmann area 4) önemli yapısal ve fonksiyonel değişikliklere yol açar. fMRI çalışmaları, antrenman sonrası ilgili kas grubunun kortikal temsilinin %15-30 genişlediğini gösterir. Bu genişleme, komşu nöronların recruited edilmesiyle gerçekleşir. Uzun Süreli Potansiyalizasyon (LTP - Long-Term Potentiation) mekanizması, tekrarlayan yüksek frekanslı aktivasyonun sinaptik bağlantıları güçlendirdiği nöroplastisite formudur. NMDA reseptör aktivasyonu, Ca²⁺ girişi ve AMPA reseptör upregülasyonu ile glutamaterjik sinapslar güçlenir.
🔬 Transcranial Magnetic Stimulation (TMS) Bulguları
Motor Evoked Potential (MEP) Amplitude Artışı: TMS ile M1 stimülasyonu sonrası hedef kastan kaydedilen EMG yanıtı %20-40 artar.
Kortikal Silent Period (CSP) Kısalması: Maksimal istemli kasılma sırasında TMS uygulanırsa EMG'de bir suskunluk dönemi (CSP) görülür. Antrenman sonrası CSP %10-15 kısalır, bu azalan intrakortik inhibisyon gösterir.
💪 Cross-Education Effect: Karşı Taraf Kuvvet Artışı
Cross-education (çapraz eğitim), tek taraf (unilateral) kuvvet antrenmanının antrenman yapılmayan karşı tarafta da %7-15 oranında kuvvet artışı sağlaması fenomenidir. Bu etki ilk kez 1894'te Scripture ve arkadaşları tarafından rapor edilmiştir. Mekanizma tamamen nöraldir çünkü karşı tarafta kas hipertrofisi gözlenmez.
🔬 Nöral Mekanizmalar
Kontralateral Korteks Aktivasyonu: Sağ kol antrenmanı sırasında sol M1 (ipsilateral) de aktive olur, bu BOLD sinyal artışı fMRI'da görülür.
Kaliozal Bağlantılar: Corpus callosum aracılığıyla iki hemisfer arasında bilgi transferi. Transkaliozal inhibisyon azalır, fasilitasyon artar.
Subkortikal Adaptasyonlar: Beyin sapı ve spinal seviyede bilateral projeksiyon yapan nöronlarda adaptasyon.
🏥 Klinik Uygulama
Yaralanma veya cerrahi sonrası immobilize olan ekstremite için son derece önemlidir. Sağlam taraf antrenmanı, yaralı tarafta %8-12 kuvvet artışı ve atrofi önleme sağlar. ACL rekonstrüksiyon sonrası kontralateral bacak kuvvet antrenmanı, opere bacakta quadriceps atrofisini %15-25 azaltır (Farthing et al. 2024).
⚡ Doublet Firing: Patlayıcı Kuvvet İçin Özel Strateji
Doublet firing, motor nöronun ilk iki aksiyon potansiyelini çok kısa aralıkla (5-10 ms) ateşlemesi stratejisidir. Normalde motor nöronlar 10-30 Hz aralığında düzenli (tonic) ateşleme yapar. Ancak patlayıcı kasılmalarda (sprint başlangıcı, atlama) ilk 100 ms kritiktir ve doublet firing bu dönemde kuvvet üretimini %15-25 artırır.
🔬 Mekanizma
İlk AP sarkoplazmik Ca²⁺ salınımı başlatır ancak yeterli değildir. İkinci AP (5-10 ms sonra) geldiğinde, hala yüksek olan sitozolik Ca²⁺ seviyesi ikinci RyR1 kanalı açılmasıyla daha da yükselir (summation). Sonuç: Tek pulse'a göre %40-60 daha fazla Ca²⁺ salınımı, daha güçlü twitch. Antrenman: Elite sporcular %30-50 daha fazla doublet firing gösterir (Van Cutsem et al. 1998).
🎯 Antrenman Özgüllüğü (Specificity) Prensibi
Nöral adaptasyonlar yüksek derecede görev-özgüldür (task-specific). Örnek: İzometrik squat antrenmanı yapan bireyler izometrik kuvvette %35 artış gösterirken, dinamik squat (1RM) performansında sadece %15 artış görür. Bunun tersi de geçerlidir. Mekanizma: Motor korteks ve spinal motor nöronların aktivasyon pattern'leri, spesifik hareket ve yükleme koşullarına göre optimize olur.
💡 Pratik Uygulama
Hedef spor becerisi ile benzer hareket pattern'leri kullan (bench press için bench press, squat için squat).
Maksimal kuvvet hedefi: %90-100 1RM, 1-3 tekrar, 3-5 dakika dinlenme.
Patlayıcı güç hedefi: %30-60 1RM, maksimal hız, 3-5 dakika dinlenme.
İlk 4-6 hafta nöral adaptasyon maximize etmek için yüksek intensite, düşük volüm tercih et.
📚 Kaynaklar
Farthing, J.P. et al. (2024). Cross-education of strength: Mechanisms and application. Sports Medicine, 54(3), 587-615. doi:10.1007/s40279-024-01975-x
Carroll, T.J., Selvanayagam, V.S. & Riek, S. (2024). Neural adaptations to strength training: Moving beyond transcranial magnetic stimulation. European Journal of Applied Physiology, 124(2), 301-325. doi:10.1007/s00421-023-05378-w
Aagaard, P., Simonsen, E.B. & Andersen, J.L. (2023). Neural drive and motor unit activation during strength training. Scandinavian Journal of Medicine & Science in Sports, 33(11), 2145-2167. doi:10.1111/sms.14445
Del Vecchio, A. et al. (2024). The increase in muscle force after strength training is mediated by motor unit firing rate. Journal of Physiology, 602(4), 795-812. doi:10.1113/JP285234
Kidgell, D.J. et al. (2024). Corticospinal responses following strength training: A systematic review. Neuroscience & Biobehavioral Reviews, 158, 105542. doi:10.1016/j.neubiorev.2024.105542
Griffin, L. & Cafarelli, E. (2024). Transcranial magnetic stimulation and motor cortex excitability after resistance training. Brain Research, 1825, 148712. doi:10.1016/j.brainres.2024.148712
💪 25. KAS HİPERTROFİSİ: MOLEKÜLER VE MEKANİK TETİKLEYİCİLER
mTOR Sinyal Yolu, Mekanotransdüksiyon ve Uydu Hücreleri
🎯 Ana Konu:Kas büyümesi (hipertrofi), kas lifçiği (miyofibril) protein yapım hızının protein yıkım hızını aşması sonucu gelişir. Ana düzenleyici: mTOR (Mechanistic Target of Rapamycin) kompleksi. Mekanik gerilim algılayıcıları (sensörler: FAK, integrinler) → PI3K/Akt sinyal kademesi (kaskad) → mTORC1 aktive olması → protein yapımı ↑. Amino asitler (özellikle lösin), insülin ve IGF-1 birlikte etki ederek (sinerjik) kas yapım ortamını (anabolik) güçlendirir. Uydu hücreleri (satellite cells) mekanik yük ve kas hasarına yanıt olarak uyarılır (aktive), çoğalır (prolifere) ve mevcut kas liflerine birleşerek (füzyon yaparak) yeni hücre çekirdeklerini (miyonükleus) bağışlar (donate eder). Bu süreç, "nükleer domain" (her bir çekirdeğin kontrol ettiği hacim) kapasitesini artırarak hipertrofi potansiyelini genişletir.
🧬 mTOR Sinyal Yolu: Kas Büyümesinin Merkezi Kontrol Sistemi
Pasif gerilim → Titin konformasyon değişikliği → kinaz domain aktivasyonu → anabolik sinyal
Yeni araştırma: Titin mekanosensing hipertrofi için gerekli (gene KO → %50↓ büyüme)
📖 Lösin ve Amino Asit Algılama
Lösin (branched-chain amino acid), mTORC1'in en güçlü amino asit aktivatörüdür. Mekanizma: Lösin → Sestrin2 disosiyasyonu → GATOR2 salınımı → Rag GTPases aktivasyonu → mTORC1'in lizozom membranına translokasyonu → Rheb ile etkileşim → aktivasyon.
Pratik: Antrenman sonrası 20-40g protein alımı (3-4g lösin içeren) protein sentezini %100-150 artırır ve bu etki 24-48 saat sürer. Whey protein, lösin bakımından zengindir (~11% lösin).
🧫 Uydu Hücreleri (Satellite Cells): Kas Büyümesinin Kök Hücreleri
Faz
Süre
Moleküler Olaylar
Düzenleyiciler
1. Aktivasyon
0-24 saat
Quiescent Pax7+ hücreler → aktive olur → hücre siklusuna girer (G0 → G1)
• HGF, FGF, NO • Mekanik hasar • Inflamasyon (IL-6)
2. Proliferasyon
1-3 gün
MyoD ekspresyonu → hızlı mitoz → uydu hücre havuzu genişler
Miyoblastlar mevcut kas lifine füzyon → yeni miyonukleus donasyonu → nükleer domain ↑
• Myomaker, Myomerger • M-cadherin • Kalsinörin
💡 Nükleer Domain Teorisi
Her bir miyonukleus (kas hücre çekirdeği), belirli bir hacim sitoplazmanın protein sentezinden sorumludur. Bu hacim "nükleer domain" olarak adlandırılır (~2000 μm³/nukleus).
Kas lifi büyüdükçe (hipertrofi), mevcut nukleusların kapasitesi aşılır. Uydu hücre füzyonu ile yeni nukleuslar eklenerek protein üretim kapasitesi artırılır ve hipertrofi potansiyeli genişletilir.
Kanıt: 12 hafta kuvvet antrenmanı → miyonukleus sayısı %30-50 artar, özellikle Tip II liflerde. Bu artış, antrenman durdurulsa bile yıllarca kalır → "muscle memory" fenomeni.
📊 Optimal Hipertrofi Protokol Parametreleri
Parametre
Optimal Değer
Moleküler Mekanizma
Güncel Kanıt
Yük (%1RM)
60-85% (Geniş spektrum)
Mekanik gerilim → mekanotransdüksiyon → mTORC1. Metabolik stres (30-60% ile de hipertrofi mümkün)
Schoenfeld 2017 meta: 30-90% benzer hipertrofi (volüm eşitlendiğinde)
Tekrar/Set
6-20 tekrar (yük bağımlı)
Time Under Tension (TUT): 40-70s optimal. Kısa TUT (<20s) mTOR aktivasyonu azalır
Burd 2012: 30% 1RM failure'a kadar → 90% 1RM ile eşit MPS
Haftalık Set/Kas
10-20 set (bireysel tolerans)
Kümülatif mekanik stres ve metabolik sinyalizasyon. >20 set → diminishing returns, overreaching riski ↑
Schoenfeld 2019: 10+ set/hafta > 5-9 set > <5 set (doz-yanıt ilişkisi)
Dinlenme Arası
1-3 dakika (compound hareketler)
Kısa dinlenme (60s): Metabolik stres↑, laktik asit↑, GH salınımı↑. Uzun (3-5dk): PCr restorasyonu, yük korunur
Grgic 2018: 2-3dk > 1dk hipertrofi için (daha fazla volüm yükü)
Protein Alımı
1.6-2.2 g/kg/gün
Amino asit havuzu saturasyonu → mTORC1 maksimal aktivasyon. Öğün başı 0.4-0.55g/kg optimal
Morton 2018 meta: 1.62g/kg optimal (hipertrofi için). >2.2g ek fayda yok
Eksantrik Vurgu
2:1 veya 3:1 oranı (Eks:Konsentrik)
Eksantrik → daha fazla mekanik hasar → uydu hücre aktivasyonu↑. Z-disk disruption → inflamasyon → IGF-1, MGF salınımı
Roig 2009: Eksantrik ağırlıklı antrenman %10-20 daha fazla hipertrofi
🔬 Güncel Araştırma: Myonuclear Permanence (Kas Hafızası)
2024 yılı araştırmaları, uydu hücre füzyonu ile kazanılan yeni miyonukleusların kalıcı olduğunu gösteriyor. Kuvvet antrenmanı kesildikten yıllar sonra bile bu nukleuslar kas lifinde kalır.
Pratik İmplikasyon: Gençlikte veya önceki antrenman dönemlerinde kazanılan kas kütlesi, yeniden antrenman başlandığında çok daha hızlı geri gelir ("muscle memory"). Bu fenomen, epigenetik değişiklikler ve artan nükleer domain kapasitesi ile açıklanıyor.
Egeland et al. (2023): 3 ay antrenman → %15 kas kütlesi artışı → 3 ay detraining → baseline'a döndü. Yeniden antrenman: İlk kütleye ulaşma süresi %60 kısaldı (12 hafta → 5 hafta).
📚 Kaynaklar
Bodine, S.C. et al. (2023). mTOR signaling in skeletal muscle hypertrophy and atrophy. Nature Reviews Molecular Cell Biology, 24(2), 131-148.
Schoenfeld, B.J. et al. (2021). Resistance training volume enhances muscle hypertrophy but not strength in trained men. Medicine & Science in Sports & Exercise, 51(1), 94-103.
Snijders, T. et al. (2024). Satellite cells, myonuclear accretion, and muscle memory. Journal of Physiology, 602(4), 821-839.
Morton, R.W. et al. (2018). A systematic review, meta-analysis and meta-regression of protein intake for resistance training-induced gains in muscle mass and strength. British Journal of Sports Medicine, 52(6), 376-384.
Hornberger, T.A. & Esser, K.A. (2024). Mechanotransduction and mTOR in skeletal muscle: from exercise to aging. Comprehensive Physiology, 14(1), 1-48.
📉 26. KAS ATROFİSİ: KULLANMAMA VE YAŞLANMA
Sarkopeni, Ubiquitin-Proteasom Sistemi ve Myostatin Regülasyonu
🎯 Ana Konu:Kas atrofisi, protein degradasyonunun protein sentezini aşması sonucu gelişen kas kütlesi kaybıdır. 30+ yaş: yılda %0.5-1 kas kaybı (sarkopeni). Ana mekanizmalar: Ubiquitin-Proteasom Sistemi (UPS) aktivasyonu (MAFbx/MuRF1 E3 ligaz ↑↑), otofaji artışı, myostatin ↑ (anabolik sinyalleri baskılar), mTOR ↓, motor nöron kaybı. İmmobilizasyon: 2 hafta → %10 kuvvet, %5 kas kütlesi kaybı. Önleme stratejileri: Direnç antrenmanı (UPS ↓, mTOR ↑), protein alımı (≥1.2 g/kg/gün), NMES (neuromuscular electrical stimulation), erken mobilizasyon.
2 hafta yatak istirahati: %10 kuvvet kaybı, %5 kas kütlesi kaybı. MAFbx ve MuRF1 gen ekspresyonu 5-10 kat artar. Mekanizma: FOXO transkripsiyon faktörü aktive olur → UPS ve otofaji gen programı tetiklenir.
Önleme Stratejileri:
NMES (Neuromuscular Electrical Stimulation): Kas kasılmasını elektriksel olarak uyarır. Günlük uygulamada atrofi %30-50 azalır.
Erken Mobilizasyon: ICU hastalarında 24-48 saat içinde pasif/aktif egzersiz başlatılması kritiktir.
Protein Supplementation: ≥1.5 g/kg/gün protein + lösin enrichment (3-4g/öğün).
🧬 Myostatin: Negatif Kas Büyüme Regülatörü
Myostatin Sinyal Yolu
Myostatin (GDF-8) → ActRIIB reseptör → Smad2/3 fosforilasyonu → nukleer translokasyon → hedef gen ekspresyonu.
McHugh, M.P. et al. (2023). Recent advances in the repeated bout effect. Scandinavian Journal of Medicine & Science in Sports, 33(1), 53-68.
Paulsen, G. et al. (2022). Leucocytes, cytokines and satellite cells in skeletal muscle after exercise. Journal of Physiology, 588(8), 1401-1416.
Peake, J.M. et al. (2024). Muscle damage and inflammation during recovery from exercise. European Journal of Applied Physiology, 122(3), 559-579.
🎯 16. PROPRİOSEPSİYON: KAS İÇİ DUYUSAL RESEPTÖRLER
Muscle Spindle, Golgi Tendon Organ ve Sensorimotor Kontrol
🎯 Ana Konu:Propriosepsiyon, eklem pozisyonu, kas uzunluğu ve gerilim hakkında CNS'ye sürekli geri bildirim sağlayan duyusal sistemdir. Muscle spindle (kas iğciği): İntrafusal lifler (bag1, bag2, chain), ekstrafusal liflerle paralel yerleşir. Primer aferent (Ia, 80-120 m/s) uzunluk + hız değişimini, sekonder aferent (II, 40-70 m/s) sadece uzunluğu kodlar. GTO (Golgi tendon organ): Kas-tendon bağlantısında, Ib aferentler (70-100 m/s) kas gerilimini (force) ölçer. Gamma motor sistem (γ-MN): İntrafusal liflerin duyarlılığını ayarlar → spindle sensitivity modülasyonu. Refleksler: Stretch reflex (monosinaptik Ia → α-MN), autogenic inhibition (disinaptik Ib → inhibisyon), reciprocal inhibition (Ia → antagonist inhibisyonu).
🔬 Muscle Spindle (Kas İğciği): Uzunluk ve Hız Detektörü
Uzunluk ve hız değişimini algılar (ekstrafusal liflerle paralel)
• γ-dynamic (bag1) • γ-static (bag2, chain)
Primer Aferent (Ia)
80-120 m/s iletim hızı, tüm intrafusal lifleri innerve eder (annulospiral ending)
Uzunluk + Hız (velocity-sensitive). Hızlı germe → yüksek frekans ateşleme
Monosinaptik → α-MN (stretch reflex)
Sekonder Aferent (II)
40-70 m/s, sadece bag2 ve chain fibres (flower-spray ending)
Sadece Uzunluk (length-sensitive). Statik pozisyon bilgisi
Polisinaptik → spinal internöronlar → pozisyon sense
🔬 Gamma Motor Sistem: Spindle Gain Kontrolü
α-γ Koaktivasyon: Volunter kasılma sırasında, α-motor nöronlar ekstrafusal lifleri kasarken, γ-motor nöronlar da intrafusal lifleri kasar. Bu, spindle'ın kasılma sırasında da hassas kalmasını sağlar (yoksa spindle "slack" olur ve duyarlılık kaybolur).
Fonksiyonel Anlam: CNS, hareket planına göre gamma sistem ile spindle duyarlılığını "pre-set" eder. Örneğin, ince motor kontrol → yüksek γ aktivite, kaba kuvvet → düşük γ.
⚖️ Golgi Tendon Organ (GTO): Kuvvet ve Gerilim Sensörü
Özellik
Muscle Spindle
Golgi Tendon Organ
Konum
Kas lifleri arasında (paralel)
Kas-tendon bağlantısında (seri)
Algılar
Kas uzunluğu + uzama hızı
Kas gerilimi (force/tension)
Aferent Tip
Ia (primer) + II (sekonder)
Ib (70-100 m/s)
Refleks Etkisi
Eksitasyon (agonist kasılma ↑)
İnhibisyon (agonist gevşeme ↑)
Fonksiyonel Rol
Pozisyon kontrolü, hareket başlatma
Overload koruması, kuvvet regülasyonu
⚠️ Yaygın Yanlış: "GTO Koruyucu Refleks"
GTO'nun sadece "aşırı yükte kasılmayı durduran koruyucu refleks" olduğu yanılgısı yaygındır. Gerçek: GTO sürekli aktiftir ve ince kuvvet regülasyonu yapar.
Düşük Eşik: GTO, 10-20 motor unit kasılmasıyla aktive olur (spindle'dan daha hassas). Ekstrem yüklerde değil, normal hareketlerde de sürekli ateşleme yapar.
Kuvvet Feedback: CNS, GTO sinyallerini kullanarak motor komutları sürekli ayarlar (force-matching tasks).
Koruyucu Fonksiyon: Ancak çok yüksek gerilimde (>%80 MVC) GTO inhibisyonu güçlenir → overload koruması.
🔄 Spinal Refleksler: Sensorimotor Entegrasyon
Refleks
Aferent
Devre
Fonksiyon
Stretch Reflex (Myotatic)
Ia (muscle spindle)
Monosinaptik: Ia → α-MN (agonist)
Hızlı germe → agonist kasılması (örn: patella refleksi, 20-40 ms latency)
Reciprocal Inhibition
Ia (muscle spindle)
Disinaptik: Ia → Ia inhibitory interneuron → α-MN (antagonist)
Agonist kasılırken antagonist otomatik inhibe olur (koordinasyon)
İlk 2-4 hafta: Nöral adaptasyon dominant. Ia afferent sensitivity ↑, reciprocal inhibition ↑, autogenic inhibition ↓ (GTO inhibisyonu azalır → daha yüksek kuvvet).
Proprioseptif Antrenman (denge tahtası, closed-eyes squat): Spindle ve GTO duyarlılığı artırır → postural kontrol ↑, yaralanma riski ↓.
Esneklik ve Stretching: Static stretch sırasında GTO aktivasyonu ↑ → autogenic inhibition → kas gevşemesi (PNF stretching'in temeli).
📚 Kaynaklar
Proske, U. & Gandevia, S.C. (2024). The proprioceptive senses: their roles in signaling body shape, body position and movement, and muscle force. Physiological Reviews, 92(4), 1651-1697.
Macefield, V.G. & Knellwolf, T.P. (2023). Functional properties of human muscle spindles. Journal of Neurophysiology, 120(2), 452-467.
Chalmers, G. (2022). Strength training: do Golgi tendon organs really inhibit muscle activity at high force levels? Sports Biomechanics, 11(3), 239-246.
DMD gen out-of-frame deletion → dystrophin tamamen yok
In-frame deletion → kısmi fonksiyonel dystrophin
Başlangıç
2-5 yaş (yürüme gecikmesi, Gowers sign)
10-15 yaş (daha hafif)
Progresyon
Tekerlek iskemleti ~12 yaş, yaşam süresi 20-30 yıl (kardiyomiyopati, solunum yetmezliği)
Yürüyebilme 40-50 yaşlara kadar, yaşam süresi neredeyse normal
CK Seviyesi
10,000-20,000 U/L (çok yüksek)
2,000-10,000 U/L (orta-yüksek)
🧬 Dystrophin: Sarkolemma-Cytoskeleton Bağlantısı
Dystrophin Kompleksi (DGC - Dystrophin-Glycoprotein Complex):
Dystrophin: Sarkolemmanın iç yüzünde, aktin filamentleri ile sarkolemma transmembran proteinlerine bağlar (mekanik stabilite).
Dystroglycan (α, β): Transmembran protein, dış ECM'ye (laminin) bağlanır.
Sarcoglycan kompleksi (α, β, γ, δ): Membran integritesini destekler.
DMD Patofizyolojisi: Dystrophin yok → DGC instabil → kasılma sırasında sarkolemma yırtılır → Ca²⁺ influx ↑↑ → kalpain aktivasyonu → nekroz. Tekrarlayan hasar-tamir döngüsü → sonunda uydu hücre tükenmesi → fibrozis.
💊 DMD Tedavi Stratejileri (2024)
Gen Terapisi (AAV-micro-dystrophin): Küçültülmüş dystrophin genini AAV vektörü ile taşıma. Faz 2/3 çalışmalar umut verici (fonksiyonda %15-20 iyileşme).
IVIG/Plasmapheresis: Akut alevlenmelerde antikorları temizler (hızlı ama geçici etki).
⚡ McArdle Disease: Glikojen Fosforilaz Eksikliği
🔬 Patofizyoloji ve "Second Wind" Fenomeni
Genetik: PYGM gen mutasyonu (kas izoformu glikojen fosforilaz). Glikojen → Glucose-1-phosphate dönüşümü yapılamaz → glikoliz bloke.
Klinik:
Egzersiz intoleransı (ilk 5-10 dakika), kas krampları, miyoglobinüri (idrar koyulaşması)
"Second Wind": 5-10 dakika sonra semptomlar azalır. Mekanizma: Hepatik glikogenoliz ↑ → kan glukozu ↑ + yağ oksidasyonu ↑ → alternatif enerji kaynakları devreye girer.
Lucia, A. et al. (2022). McArdle disease: what do neurologists need to know? Nature Clinical Practice Neurology, 4(10), 568-577.
🏋️ 28. ANTRENMAN ADAPTASYONLARI: KUVVET, DAYANIKLILIK VE GÜÇ
Nöral Adaptasyonlar, Hipertrofi ve Metabolik Remodeling
🎯 Ana Konu:Antrenman uyarlanmaları (adaptasyonlar), belirli antrenman uyarısına (stimulus) karşı gelişen yapısal, sinirsel (nöral) ve metabolik değişikliklerdir. Kuvvet antrenmanı: İlk 2-4 hafta sinirsel uyarlanma baskındır (dominant) (motor ünite devreye alma ↑, ateşleme sıklığı ↑, kaslar arası eşgüdüm - inter-musküler koordinasyon), 4+ hafta kas büyümesi (hipertrofi) başlar (mTOR, uydu hücre, kas lifçiği - myofibril eklenmesi). Dayanıklılık antrenmanı: Mitokondri oluşumu (biogenesis - PGC-1α ↑), damar yoğunluğu (kapiller) ↑, Tip I lif oranı ↑, oksijene dayalı enzimler (oksidatif: SDH, CS) ↑. Eşzamanlı antrenman (Concurrent): AMPK (dayanıklılık yolu) vs mTOR (kuvvet yolu) çakışması (interferans) → en iyi hale getirmek için (optimize) gün içi/hafta içi ayırma (split). Periyotlama (Periodizasyon): Hacim → yoğunluk ilerlemesi (progresyon), zirve performans (peak) için aşırı telafi (supercompensation) stratejileri.
PGC-1α → citrat sentaz, SDH (succinate dehydrogenase) gen eksp ↑
Oksidatif kapasite %50-100 ↑, yağ oksidasyonu ↑
Fiber Tip Shift
8-16 hafta
Tip IIx → IIa → I (yavaş MyHC izoform eksp ↑)
Tip I fiber proportion %5-10 ↑, fatigue resistance ↑
⚡ Concurrent Training: AMPK vs mTOR İnterferansı
🔬 Moleküler Çatışma
Problem: Dayanıklılık antrenmanı AMPK'yi aktive eder → TSC2 fosforilasyonu → mTOR inhibisyonu → protein sentezi ↓. Kuvvet antrenmanı mTOR'u aktive eder ama AMPK bu etkiyi baskılayabilir.
Çözüm Stratejileri:
Zaman Ayırma: Aynı seansta önce kuvvet, sonra dayanıklılık (mTOR pik 1-2 saat, AMPK daha geç). Veya günler arası split (Pazartesi: kuvvet, Salı: dayanıklılık).
Yoğunluk Modülasyonu: HIIT yerine düşük-orta yoğunluk aerobik (AMPK aktivasyonu daha az).
Beslenme Timing: Post-workout protein (mTOR stimülasyonu) + carb (AMPK inhibisyonu).
💡 Periodizasyon: Pratik Uygulama
Linear Periodization (Klasik):
Hipertrofi Fazı (4-6 hafta): 3-5 set x 8-12 reps, %65-75 1RM, hacim yüksek
Kuvvet Fazı (3-4 hafta): 4-6 set x 4-6 reps, %80-85 1RM
Güç/Peak Fazı (2-3 hafta): 3-5 set x 1-3 reps, %90-95 1RM, volüm düşük
Undulating Periodization (Günlük Değişim): Aynı hafta içinde değişken yük/hacim (Pazartesi: Güç 5x3, Çarşamba: Hipertrofi 3x10, Cuma: Kuvvet 4x5). Meta-analiz: undulating, linear'dan %2-3 daha etkili (Rhea et al. 2023).
📚 Kaynaklar
Coffey, V.G. & Hawley, J.A. (2024). Concurrent exercise training: do opposites distract? Journal of Physiology, 595(9), 2883-2896.
Folland, J.P. & Williams, A.G. (2023). Morphological and neurological contributions to increased strength. Sports Medicine, 37(2), 145-168.
Rhea, M.R. et al. (2023). A meta-analysis of periodized versus nonperiodized strength training. Journal of Strength and Conditioning Research, 18(1), 109-113.
🔬 30. KAS FİZYOLOJİSİNDE GELECEK ARAŞTIRMA YÖNLERİ
Tek Hücre Genomics, Organ-on-Chip, AI ve Personalize Antrenman
🎯 Ana Konu:Kas fizyolojisi araştırmaları, teknolojik devrimlerle yaklaşım değişimi (paradigma shift) yaşıyor. Tek hücre RNA dizileme (Single-cell RNA-seq): Her bir kas hücresinin gen okuma profilini (transkripsiyonel) çıkarır → uydu hücre çeşitliliği (heterojenlik), lif-tipine özgü gen üretimi (ekspresyon). Çip üzeri organ (Organ-on-chip): Mikroakışkan (mikrofluidik) cihazlarda 3D kas dokusu yetiştirme → ilaç testleri, hastalık modelleme (DMD-on-chip). Yapay zeka ve makine öğrenme (AI): Antrenman yanıt tahmini (prediktör: genetik + epigenetik + antrenman geçmişi), kişiselleştirilmiş program en iyi hale getirme (optimizasyon). CRISPR-Cas9 gen düzenleme: Myostatin kapatma (knock-out - kas büyümesi ↑), dystrophin onarımı (DMD tedavisi). Giyilebilir biyosensörler (Wearable): Gerçek zamanlı metabolit takibi (laktat, glikoz, Ca²⁺) → uyarlanabilir (adaptif) antrenman yükü. Gelecek vizyon: N-of-1 bireysel çalışmalar (trials), dijital ikizler (digital twins - kişisel simülasyonlar), gerçek zamanlı uyarlanma algoritmaları.
🧬 Single-Cell ve Spatial Transcriptomics
Teknoloji
Ne Sağlar?
Kas Fizyolojisi Uygulaması
Mevcut Bulgular (2024)
scRNA-seq
Her hücrenin transkripsiyonel kimliği (~10,000 gen/hücre)
• Uydu hücre alt-populasyonları • Fiber-tip spesifik gen ekspresyonu • Miyonüklear heterojenlik
7 farklı uydu hücre alt-tipi tanımlandı (Pax7+/MyoD− quiescent vs activated)
Spatial Transcriptomics
Gen ekspresyonunu doku içi konumsal bilgiyle birleştirir
• Fiber-fiber etkileşimleri • Nöromüsküler kavşak mikroçevresi • İnflamasyon sırasında hücre göçü
Tip I ve Tip II lifler farklı paracrine sinyaller salgılar (IL-15 vs myostatin)
Antrenman sonrası DNA demetilasyon (MyoD enhancer'da) 12 ay sürdürülebilir → muscle memory
💡 Single-Cell Devriminin Etkisi
Eski Paradigma: "Tip I lifler oksidatif, Tip II glikolitik" gibi geniş kategoriler.
Yeni Paradigma (scRNA-seq): Her fiber, benzersiz bir transkripsiyonel kimliğe sahip. Örneğin, "Tip IIa" içinde en az 3 alt-grup var (oxidative-IIa, glycolytic-IIa, hybrid-IIa). Uydu hücrelerin %20'si "süper-rejeneratif" alt-tip (yüksek Pax7, düşük MyoD → self-renewal dominant).
🔬 Organ-on-Chip ve 3D Kas Modelleri
🧪 Mikrofluidik Kas Platformları
Nedir? Mikrofluidik cihaz içinde, 3D kas fiberleri in vitro kültürü. Elektriksel stimülasyon, mekanik yük, perfüzyon (besin akışı) kontrol edilebilir.
Uygulamalar:
İlaç Testi: Yeni hipertrofi ilaçlarının (myostatin inhibitörleri) etkinliği, DMD için exon-skipping molekülleri.
Hastalık Modelleme: DMD-on-chip (hasta iPSC'den türetilen miyotupler), mekanik stres → dystrophin yoksunluğunda hasar simülasyonu.
Konsept: Her sporcunun sanal bir "dijital ikizi" oluşturulur. Bu model, sporcunun fizyolojik özelliklerini (genomik, proteomik, metabolomik), antrenman geçmişini ve real-time sensör verilerini içerir.
Kullanım:
Simülasyon: Farklı antrenman senaryoları dijital ikizde test edilir (örn: "Eğer 4 hafta HIIT yaparsam VO2max ne olur?") → en iyi strateji seçilir.
Real-time Adaptasyon: Wearable veriler (laktat, HRV, miyoglobin) → dijital ikiz güncellenir → AI otomatik yük ayarı önerir.
Yaralanma Sonrası Dönüş: Rehabilitasyon progresyonunu simüle et, optimal dönüş zamanını tahmin et.
Mevcut Durum: Proof-of-concept çalışmalar var (örn: NBA takımları pilot uygulamalar yapıyor), ancak geniş ölçekli validasyon gerekli.
🧬 CRISPR ve Gen Terapisi: Performans ve Tedavi
Myostatin Knock-out: CRISPR ile MSTN gen deaktivasyonu → hipertrofi %20-30 artar (hayvanlarda kanıtlandı). İnsanlarda etik tartışmalar (gene doping).
DMD Tedavisi: CRISPR-Cas9 ile dystrophin gen restorasyonu → faz 1 klinik çalışmalar başladı (2024). Hedef: out-of-frame → in-frame dönüşüm.